Пухоеды и вши
| Пухоеды и вши | |
|---|---|
 Лобковая вошь (Pthirus pubis) | |
| Научная классификация | |
Домен: Эукариоты Царство: Животные Подцарство: Эуметазои Без ранга: Двусторонне-симметричные Без ранга: Первичноротые Без ранга: Линяющие Без ранга: Тип: Членистоногие Подтип: Трахейнодышащие Надкласс: Шестиногие Класс: Насекомые Подкласс: Крылатые насекомые Инфракласс: Новокрылые Клада: Надотряд: Psocodea Отряд: Пухоеды и вши | |
| Международное научное название | |
| Phthiraptera Haeckel, 1896 | |
| Подотряды | |
| |
Пухое́ды и вши (лат. Phthiraptera — др.-греч. φθείρ — вошь и ἄπτερος — бескрылый) — отряд паразитических насекомых с неполным превращением. Phthiraptera многочисленны и космополитны, паразитируют, как предполагается, на всех видах птиц и на очень многих видах млекопитающих. В настоящее время учёными описано 5136 видов, включая 1 ископаемый вид (Zhang, 2013)[1]. Основу многообразия составляют два подотряда пухоедов: Amblycera (1360 описанных видов) и Ischnocera (3080 видов)[2]. Третий подотряд пухоедов — Rhynchophthirina содержит всего три вида. Последняя группа — это собственно вши (Anoplura, 543 вида), к которым относятся и три паразита человека: головная, платяная и лобковая вши.
Морфология
Пухоеды и сосущие вши — мелкие бескрылые насекомые, длиной от 0,8—11 мм[3]. Тело уплощено дорзовентрально. У пухоедов Amblycera и Ischnocera голова широкая, шире груди. В группах Rhynchophthirina и Anoplura, напротив, голова уже груди. В зависимости от локализации и общей стратегии поведения пухоеды подразделяются на несколько групп, различающихся формой тела. Так, например, среди птичьих Ischnocera можно выделить вытянутые «крыловые» и компактные «головные» и «туловищные» формы. Грудь более или менее слитная. Полное слияние наблюдается у Anoplura; у пухоедов Rhynchophthirina и Ischnocera слиты мезо- и метанотум, у Amblycera же между ними имеется шов. Брюшко состоит из 8-10 видимых сегментов. Грудь несёт одну пару отверстий трахейной системы, брюшко — от одной до шести пар.
Глаза имеются лишь у некоторых видов, и даже в этих случаях — редуцированные; простые глазки редуцированы полностью. Антенны короткие, включают до 5 члеников. У Amblycera антенны спрятаны в ямках по бокам головы, состоят из 4 члеников, третий членик стебельковидный. У остальных групп антенны свободные, нитевидные, состоят из 3-5 члеников. У некоторых пухоедов усики диморфны: принимают участие в захвате самки самцом во время спаривания[3].
Ротовые придатки пухоедов грызущего типа представлены развитыми верхней губой, мандибулами и уменьшенными максиллами и нижней губой. Максиллярный щупик сохраняется только у Amblycera. Эта же группа пухоедов выделяется и тем, что ротовые придатки у них двигаются в вертикальной плоскости — перпендикулярно оси головы, у Ischnocera, напротив, эта плоскость параллельна голове. У Rhynchophthirina ротовой аппарат тоже грызущего типа, но расположен на конце длинного выроста головы — хоботка — и повёрнут на 180°.
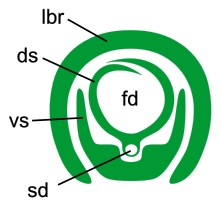
В отличие от пухоедов, все сосущие вши (Anoplura) имеют сильно видоизменённый колюще-сосущий ротовой аппарат. Он состоит из двух стилетов: дорзального и вентрального, — окружённых хоботком, который к тому же армирован «зубами». Вентральный стилет, как предполагается, образован нижней губой. Происхождение дорзального стилета гораздо более дискуссионно. По распространённой версии этот стилет образован гипофаринксом, дорзальная вогнутая часть которого образует пищевую трубку. С брюшной стороны гипофаринкса проходит слюнной канал, который часто на срезах может быть отделён от основной части гипофаринкса, представляя артефактный «средний стилет». Мандибулы и, вероятно, максиллы у сосущих вшей редуцированы[4][5][6].
Крылья или их рудименты полностью отсутствуют. Ноги развиты хорошо, являются основными прикрепительными органами как пухоедов, так и сосущих вшей. У пухоедов лапки одно- или двухчлениковые, у вшей — целиком слитные. На конце лапки несут по два коготка у всех паразитов птиц и по одному коготку у большинства паразитов млекопитающих (в том числе все Anoplura). Сосущие вши цепляются за отдельные волоски, зажимая их в щели, образуемой тарзальным коготком и шипом на голени. Таким образом, образующийся просвет этого зажима должен соответствовать толщине волос хозяина, что определяет, видимо, узкую специализацию этих вшей.
Внутреннее строение типичное для насекомых. Значительный объём занимает пищеварительная система. У форм, не питающихся кровью, имеется развитый зоб, принимающий участие в накоплении и измельчении пищи. У гематофагов зоб устроен проще или отсутствует. Мальпигиевых сосудов лишь две пары.
Образ жизни
Все современные пухоеды и вши обитают на поверхности тела птиц и млекопитающих. Основная часть видов подотрядов Amblycera и Ischnocera представлена паразитами птиц, и лишь 12 % паразитирует на млекопитающих[3]. Напротив, все три вида Rhynchophthirina и виды подотряда Anoplura приурочены исключительно к млекопитающим. Подавляющее большинство из них являются паразитами и потребляют в пищу перья, отмершую кожу, кровь и кожные выделения. Во всей группе только собственно вши (Anoplura) и могут сосать кровь, представители других подотрядов являются «жующими» формами. Впрочем, очень немногие пухоеды (некоторые Amblycera, все Rhynchophthirina) тоже являются гематофагами, но получают кровь посредством своих грызущих ротовых аппаратов.
Помимо эктопаразитов встречаются необычные виды, которые занимаются поеданием яиц и нимф кожных паразитов — клещей и других Phthiraptera, в том числе и особей своего вида. Определённую, но пока неизвестную роль имеют в пищевом рационе пухоедов бактерии и грибы.
Весь жизненный цикл пухоедов и вшей в норме протекает на поверхности одной особи хозяина. Смена особи хозяина возможна в большинстве случаев только при близком контакте хозяев. Лишь некоторые Amblycera достаточно подвижны и могут активно покидать своего хозяина в случае его смерти или ухудшения здоровья. Однако и Ischnocera также могут менять хозяина, причём довольно своеобразно. На нескольких представителях было замечено, что эти пухоеды цепляются за тела мух-кровососок, которые паразитируют на тех же хозяевах. Эти мухи гораздо более подвижны и чаще меняют хозяев, но в то же время и менее специфичны в отношении выбора вида хозяина[3][7].
Жизненный цикл и репродуктивная биология
Взрослые насекомые раздельнополы, однако самки очень часто численно превосходят самцов. У некоторых Ischnocera самцы встречаются редко или вообще не обнаружены, и т.о. эти виды являются партеногенетическими. Определение пола предполагается хромосомное, но половые хромосомы внешне не дифференцированы.
Оплодотворение внутреннее, сперматофорное. Сперматофор может расходоваться самкой длительное время, пока не будет заменён при следующем спаривании. У самок нет выраженного яйцеклада, но имеются пальцевидные гоноподы, принимающие участие в откладке. Яйца (гниды) крупные, откладываются по 1—10 в день. Определённая забота о потомстве проявляется в том, что самка прикрепляет яйца железистым цементом в основания волос и перьев, часто в трудно доступных для хозяина областях. Из яйца выходит нимфа первого возраста. Нимфа с интервалами от 3 до 12 дней претерпевает последовательные линьки, переходя во второй и третий возраста. У некоторых видов нимфы выглядят внешне, как имаго, в других же случаях различия гораздо более значительны. Нимфа третьего возраста без метаморфоза линяет на имаго. Взрослые стадии живут около месяца.
Генетика
Число хромосом у вшей и пухоедов незначительно варьирует: диплоидные числа колеблются от 10 до 16[8].
Симбиотические бактерии
Wolbachia
Вольбахии (Alphaproteobacteria: Rickettsiaceae) — облигатные внутриклеточные симбиотические бактерии, родственные риккетсиям, передающиеся трансовариально (через яйца). Эти бактерии ответственны за многие репродуктивные аномалии у членистоногих, а для некоторых насекомых и нематод-филярий являются необходимыми для развития и жизнедеятельности[9].
Все исследованные виды Phthiraptera (представители трёх подотрядов) несли вольбахий. Наиболее часто встречаемые супергруппы (клады) — это обычные вольбахии членистоногих — A и B, но также встречается редкая супергруппа F. Один и тот же вид может нести от одной до четырёх линий вольбахий, в том числе и из разных супергрупп. Эффект, оказываемый вольбахиями на вшей, неизвестен, однако предполагается, что именно они вызывают преобладание самок в популяциях, а также, возможно, ответственны за появление партеногенетических видов[7][10][11].
«Вертикальный» способ передачи вольбахий должен приводить к соответствию эволюции бактерий и их хозяев. Этого, однако, никогда не наблюдается, и, очевидно, существуют способы и горизонтальной передачи. Для вшей это также справедливо, и здесь особенно загадочен вопрос о «переносчиках» вольбахий между ними. Предполагается, что у некоторых птичьих пухоедов определённую роль здесь могут выполнять мухи-кровососки — другие эктопаразиты птиц, с которыми пухоеды входят в непосредственный контакт[7].
Riesia
'Candidatus Riesia' (Gammaproteobacteria: Enterobacteriaceae) — облигатные эндосимбиотические бактерии вшей, родственники которых также являются внутренними симбионтами насекомых, передаются трансовариально. Эти бактерии обнаружены у всех изученных видов Anoplura и, видимо, не встречаются у других вшей и вообще насекомых. Подобно вольбахиям филярий Riesia являются первичными симбионтами, то есть их наличие является необходимым для жизни хозяина. Предполагается, что ризии поставляют вшам некоторые незаменимые вещества, которых, очевидно, не хватает в крови хозяев вшей.
Локализуются Riesia внутриклеточно в т. н. «желудочных дисках». В отличие от вольбахий для достижения ооцитов эти бактерии вынуждены совершать сложные миграции, включающие экстраклеточные стадии[12][13].
Симбионты пухоедов
Многие пухоеды так же, как и сосущие вши, несут первичных симбионтов. Бактерии пухоедов, однако, не имеют прямого родства с симбионтами сосущих вшей и имеют другую локализацию в теле хозяина. Симбионты пухоеда Columbicola columbae (Ischnocera) относятся к тому же семейству, что и Riesia (Gammaproteobacteria: Enterobacteriaceae), но их ближайшими родственниками являются эндосимбионты мухи-цеце и зерновых долгоносиков. У нимф и самцов эти бактерии располагаются в бактериоцитах по бокам тела, то есть не связаны с пищеварительной трубкой. Передача бактерий происходит от матери её потомкам, и потому в теле самки они вынуждены в каждом поколении хозяев мигрировать в половые трубки для достижения ооцитов[3][14].
Значение этих бактерий для пухоедов неизвестно. Предполагается, что они так же, как Riesia, участвуют в выработке веществ, отсутствующих в диете пухоедов, но необходимых им для жизни.
Патогенность
Пухоеды и вши в норме не являются летальными паразитами. Даже предполагается, что многие пухоеды птиц приносят минимальный ущерб своим хозяевам. Тем не менее в большинстве случаев отрицательный эффект может быть выявлен. Часто в основе этого эффекта лежит раздражение, вызывающее беспокойство и расчёсывания, что приводит косвенно к снижению общей жизнеспособности. Так, например, увеличение времени, необходимого для груминга[англ.] требует затрат времени и энергии, что, кроме прочего, приводит к повышению доступности для хищников более заражённых особей[3][15].
Кожные заболевания часто также являются следствием расчёсывания, но могут быть вызваны и самими насекомыми (вши человека в том числе). Некоторые из этих заболеваний являются смертельными для молодых особей хозяина.
Для перьевых паразитов предполагается, что вызванные ими изменения формы перьев могут приводить к ухудшению аэродинамических характеристик, а также к ухудшениям брачного оперения самцов. Те формы, что поедают пух приводят косвенно к истощению, поскольку заражённой птице приходится тратить больше энергии на согревание тела[3].
Другой, часто более опасной стороной паразитизма пухоедов и вшей является факт их роли в качестве переносчиков возбудителей опасных болезней хозяев. К этим возбудителям относятся бактерии, круглые и плоские черви, грибы, а также, вероятно, и вирусы[3][16]. Среди вшей человека все три вида (Pediculus humanus, P. capitis и Pthirus pubis) могут являться переносчиками опасных бактериальных возбудителей, однако лишь Pediculus humanus в человеческих популяциях играет такую роль. Этот вид переносит три бактериальных агента: Rickettsia prowazekii, Borrelia recurrentis и Bartonella quintana[17][18].
Rickettsia prowazekii
Rickettsia prowazekii (Alphaproteobacteria: Rickettsiaceae) — возбудитель эпидемического сыпного тифа. Риккетсии заражают вшей перорально и затем поражают клетки эпителия средней кишки. Здесь они активно размножаются, а после — выходят в просвет кишки. Заражение человека происходит фекалиями вшей, попадающими в ранки на коже при расчёсывании укушенных мест. Риккетсии являются смертельными для вшей, вызывая разрыв стенки кишки и смешение содержимого кишки и гемолимфы («красные вши»). В настоящее время заболевание является редким, бактерий находят преимущественно у бездомных. Неблагоприятная ситуация остаётся в Бурунди и среди бездомных, по крайней мере, в России.
Borrelia recurrentis
Borrelia recurrentis (Spirochaetes: Spirochaetaceae) — возбудитель возвратного тифа. Вши заражаются при питании. Боррелии проникают через стенку кишки в гемоцель, где и задерживаются. Заражение человека происходит при раздавливании вши человеком, когда содержимое гемолимфы попадает на повреждённые участки кожи. В настоящее время заболевание локализовано в Эфиопии.
Bartonella quintana
Bartonella quintana (Alphaproteobacteria: Bartonellaceae) — возбудитель окопной лихорадки. Жизненный цикл сходен с таковым Rickettsia prowazekii, однако бартонелла размножается не в эпителии, а в просвете кишки, и для вшей эти бактерии не смертельны. Заражение человека происходит так же — с фекалиями паразита. Bartonella обнаружена во вшах по всему свету.
Происхождение
Происхождение Phthiraptera связывают с группой сеноедов (Psocoptera). В пользу этого предположения свидетельствует целый ряд несомненных морфологических синапоморфий. Однако основные споры ведутся относительно исторических взаимоотношений этих двух групп: являются ли они сестринскими или вши произошли от сеноедов. Кладистический анализ морфологических и молекулярных данных свидетельствует в пользу второго варианта. В соответствии с этими сценариями одно из семейств сеноедов — Liposcelidae — является сестринским по отношению ко всем Phthiraptera[3]. Недавние исследования с привлечением такого популярного гена в кладистических реконструкциях, как ядерный ген рРНК малой субъединицы рибосомы, показал неожиданные результаты. Liposcelidae и ещё одно семейство сеноедов оказываются сестринскими по отношению к Amblycera, а вся эта клада — сестринской по отношению к другим пухоедам (Anoplura в том числе). Таким образом, пухоеды возникают в пределах одной клады сеноедов, но независимо — из двух близких «источников»[19].
Вероятный сценарий перехода к паразитизму в этой ветви (один раз или дважды) включает эволюцию от комменсальных форм к облигатным паразитам. Так, многие Psocoptera (среди них — и некоторые Liposcelidae) могут обитать на волосяном и перьевом покровах и в гнёздах теплокровных позвоночных. Вопрос о первичности млекопитающих или птиц как хозяев вшей остаётся пока предметом спекуляций.
В ископаемом состоянии вши хорошо известны с плейстоценовых млекопитающих из вечной мерзлоты. В балтийском янтаре также находят волосы с гнидами. Самая древняя достоверная находка телесной Phthiraptera — это эоценовая амблицера Megamenopon rasnitsyni. Из раннего мела Забайкалья также известно насекомое Saurodectes[англ.], внешне напоминающее вошь. Его облик говорит об эктопаразитическом образе жизни, однако огромные для вшей размеры (17 мм) и некоторые другие черты говорят против фтираптероидной гипотезы[20][21].
Из-за фрагментарности и низкой достоверности древних останков время возникновения Phthiraptera пытаются определить по косвенным данным. Разные методы дают значительно расходящиеся результаты: от позднего мела до позднего карбона[].
В 2019 году из мелового периода были описаны нимфы Mesophthirus engeli из семейства Mesophthiridae, близкого к пухоедам и вшам[22].
Филогения
Относительно давно появились представления о том, что вши произошли от пухоедов, однако ясность в исторических отношениях подотрядов внесли кладистические реконструкции.
Голофилия отдельных подотрядов Phthiraptera подтверждается хорошими морфологическими апоморфиями (сомнения остаются относительно огромной группы Ischnocera). Это же подтверждают и работы по молекулярным признакам[2][23][24][25][26].
Самой базальной группой являются Amblycera. Крону составляют Rhynchophthirina и Anoplura, а Ischnocera является сестринской группой этой последней кладе. При учёте последних достижений в области реконструкции истории всей группы Psocodea эта общая схема взаимоотношений групп пухоедов и вшей принципиально не изменяется (см. выше).
| Phthiraptera |
| ||||||||||||||||||
Ограниченная подвижность, почти строгая приуроченность к определённой особи хозяина, а для вшей — и соответствие органов прикрепления толщине волос хозяина являются предпосылками для координированной эволюции паразитов и их хозяев — к их коэволюции. Косвенным свидетельством коэволюции является факт распространённой специфичности пухоедов и вшей в отношении выбора хозяев: многие виды заражают лишь один вид хозяев. Однако исключения столь же поразительны: хозяевами Menacanthus eurysternus являются 118 видов птиц из 20 семейств[3]! На уровне семейств специфичность часто проявляется в выборе хозяев из одной или нескольких групп (см. ниже). Строгий анализ соответствия кладограмм этих паразитов кладограммам их хозяев показал, что значительную долю разнообразия вшей и пухоедов на уровне родов можно объяснить коэволюцией. Кроме того, заметную роль играют такие явления, как смена хозяев и независимое видообразование у паразита[27][28][29].
Система
Система Phthiraptera ранее представляла собой два отдельных отряда: Mallophaga (пухоеды) и Anoplura (сосущие вши). Стремление к отражению филогении в системе приводило к понижению рангов этих двух отрядов до подотрядов в пределах Phthiraptera, а позднее и к уравниванию рангов всех клад пухоедов и вшей. Последнюю ситуацию мы и наблюдаем сегодня, имея четыре подотряда: Amblycera, Ischnocera, Rhynchophthirina и Anoplura. Система семейств внутри подотрядов является довольно стабильной. Однако взгляды на систему Ischnocera не устоялись: некоторые семейства и даже рода предполагаются пара- или полифилетическими, а подавляющее большинство видов сосредоточено в сборном семействе Philopteridae[24].
Семейства отряда Phthiraptera (в скобках указан круг хозяев)[20][24]:
1. Подотряд Amblycera
- Boopiidae (австралийские сумчатые)
- Gyropidae (грызуны)
- Laemobothriidae (пастушковые, аистовые, ястребиные)
- Menoponidae (птицы)
- Ricinidae (колибри, воробьинообразные)
- Trimenoponidae (южноамериканские сумчатые, грызуны)
2. Подотряд Ischnocera
- Goniodidae (птицы)
- Heptapsogasteridae (птицы)
- «Philopteridae» (птицы) (Harrisoniella hopkinsi)
- Trichodectidae (плацентарные)
- Trichophilopteridae (лемуровые, индриевые)
3. Подотряд Rhynchophthirina
- Haematomyzidae (слоны, два вида свиней)
4. Подотряд Anoplura
- Echinophthiriidae (ластоногие, куньи)
- Enderleinellidae (беличьи)
- Haematopinidae (копытные)
- Hamophthiriidae (шерстокрылые)
- Hoplopleuridae (насекомоядные, грызуны, зайцеобразные, приматы)
- Hybothiridae (трубкозуб)
- Linognathidae (копытные, псовые, даманы)
- Microthoraciidae (верблюдовые)
- Neolinognathidae (прыгунчики)
- Pecaroecidae (пекариевые)
- Pedicinidae (мартышковые обезьяны)
- Pediculidae (цепкохвостые и человекообразные обезьяны)
- Pthiridae (человекообразные обезьяны)
- Polyplacidae (грызуны, насекомоядные, приматы, зайцеобразные)
- Ratemiidae (лошадиные)
Охранный статус
Несмотря на преимущественно негативное взаимодействие вшей с человеком, 1 их вид занесён в Красный список угрожаемых видов МСОП (The IUCN Red List of Threatened Species) в категорию на грани исчезновения (CR)[30]:
 Haematopinus oliveri — находящийся на грани исчезновения вид вшей из семейства Haematopinidae.
Haematopinus oliveri — находящийся на грани исчезновения вид вшей из семейства Haematopinidae.
Примечания
- ↑ Zhang, Z.-Q. «Phylum Athropoda». — In: Zhang, Z.-Q. (Ed.) «Animal Biodiversity: An Outline of Higher-level Classification and Survey of Taxonomic Richness (Addenda 2013)». (англ.) // Zootaxa / Zhang, Z.-Q. (Chief Editor & Founder). — Auckland: Magnolia Press, 2013. — Vol. 3703, no. 1. — P. 17—26. — ISBN 978-1-77557-248-0 (paperback) ISBN 978-1-77557-249-7 (online edition). — ISSN 1175-5326. Архивировано 19 сентября 2013 года.
- ↑ 1 2 Smith, V.S. (2003). Lousy phylogenies: Phthiraptera systematics and the antiquity of lice. Proceedings of 1st Dresden Meeting on Insect Phylogeny: 150—151
- ↑ 1 2 3 4 5 6 7 8 9 10 Johnson, K.P. & D.H. Clayton (2003). The biology, ecology, and evolution of chewing lice. In: The chewing lice: World checklist and biological overview. Illinois Natural History Survey Special publication 24: 1-25
- ↑ Kristensen, N.P. (2005). A compendum of hexapod systematics. Natural History Museum of Denmark, University of Copenhagen. 44 pp.+figs.
- ↑ Fernando, W. (1933). The development and homologies of the mouth-parts of the head-louse. Quarternary Journal of Microscopic Sciences, New Series 76: 231—241
- ↑ Gullan, J.P., P.S. Cranston & K. Hansen McInnes (2004). The insects: an outline of entomology. Blackwell Publ. 505 pp.
- ↑ 1 2 3 Covacin, C. & S.C. Barker (2007). Supergroup F Wolbachia bacteria parasitise lice (Insecta: Phthiraptera). Parasitology Research 100 (3): 479—485
- ↑ Blackmon, Heath. Synthesis of cytogenetic data in Insecta // Synthesis and phylogenetic comparative analyses of the causes and consequences of karyotype evolution in arthropods (англ.). — Arlington: The University of Texas at Arlington, 2015. — P. 1—26. — 447 p. — (Degree of Doctor of Philosophy). Архивировано 14 сентября 2022 года.
- ↑ Werren, J.H. (1997). Biology of Wolbachia. Annual Reviews — Entomology 42: 587—609
- ↑ Perotti, M.A., S.S. Catalá, A.V. Ormeño, M. Żelazowska, S.M. Biliński & H.R. Braig (2004). The sex ratio distortion in the human head louse is conserved over time. Genetics 5 (10): 1-13
- ↑ Kyei-Poku, G.K., D.D. Colwell, P. Coghlin, B. Benkel & K.D. Floate (2005) On the ubiquity and phylogeny of Wolbachia in lice. Molecular Ecology 14 (1): 285—294
- ↑ Sasaki-Fukatsu, K., R. Koga, N. Nikoh, K. Yoshizawa, S. Kasai, M. Mihara, M. Kobayashi, T. Tomita & T. Fukatsu (2006). Symbiotic bacteria associated with stomach discs of human lice. Applied and Enviromental Microbiology 72 (11): 7349-7352
- ↑ Allen, J.M., D.L. Reed, M.A. Perotti & H.R. Braig (2007). Evolutionary relationships of Candidatus Riesia spp., endosymbiotic Enterobacteriaceae living within hematophagous primate lice. Applied and Enviromental Microbiology 73 (5): 1659—1664
- ↑ Fukatsu, T., R. Koga, W.A. Smith, K. Tanaka, N. Nikoh, K. Sasaki-Fukatsu, K. Yoshizawa, C. Dale & D.H. Clayton (2007). Bacterial endosymbiont of the slender pigeon louse, Columbicola columbae, allied to endosymbionts of grain weevils and tsetse flies. Applied and Enviromental Microbiology 73 (20): 6660-6668
- ↑ Kettle, D.S. (1984). Medical and veterinary entomology. Routledge Publ. 658 pp.
- ↑ Cole, 1999
- ↑ Roux, V. & D. Raoult (1999). Body lice as tools for diagnosis and surveillance of reemerging diseases. Journal of Clinical Microbiology 37 (3): 596—599
- ↑ Badiaga, S., P. Brouqui & D. Raoult (2005). Autochthonous epidemic typhus associated with Bartonella quintana bacteremia in a homeless person. The American Journal of Tropical Medicine and Hygiene 72 (5): 638—639
- ↑ Johnson, K.P., K. Yoshizawa & V.S. Smith (2004). Multiple origins of parasitism in lice. Proceedings of the Royal Society of London B 271: 1771—1776
- ↑ 1 2 Grimaldi, D.A. & M.S. Engel (2005). Evolution of the insects. Cambridge University Press. 755 pp.
- ↑ Rasnitsyn, A. P.; Zherikhin, V. V. First fossil chewing louse from the Lower Cretaceous of Baissa, Transbaikalia (Insecta, Pediculida = Phthiriaptera, Saurodectidae fam. n) (англ.) // Russian Entomological Journal : journal. — 2000. — Vol. 8. — P. 253—255. Архивировано 11 ноября 2011 года.
- ↑ Taiping Gao, Xiangchu Yin, Chungkun Shih, Alexandr P. Rasnitsyn, Xing Xu, Sha Chen, Chen Wang & Dong Ren. New insects feeding on dinosaur feathers in mid-Cretaceous amber (англ.) // Nature Communications : Журнал. — США, 2019. — Vol. 10, Article number: 5424. — P. 1—7. — doi:10.1038/s41467-019-13516-4. Архивировано 13 декабря 2019 года.
- ↑ Johnson, K.P. & M.F. Whiting (2002). Multiple genes and the monophyly of Ischnocera (Insecta: Phthiraptera). Phylogenetics and Evolution 22 (1): 101—110
- ↑ 1 2 3 Smith, V.S. (2001). Avian louse phylogeny (Phthiraptera: Ischnocera): a cladistic study based on morphology. Zoological Journal of the Linnean Society 132: 81-144
- ↑ Smith, V.S., R.D.M. Page & K.P. Johnson (2004). Data incongruence and the problem of avian louse phylogeny. Zoologica Scripta 33 (3): 239—259
- ↑ Barker, S.C., M. Whiting, K.P. Johnson, A. Murrell (2003). Phylogeny of the lice (Insecta, Phthiraptera) inferred from small subunit rRNA. Zoologica Scripta 32 (5): 407—414
- ↑ Marshall, I.K. (2002). Congruence and cospeciation: morphological and molecular phylogenetics of the Amblycera (Phthiraptera). PhD Thesis, University of Glasgow, 252 pp.
- ↑ Reed et al., 2007
- ↑ Paterson, A.M., G.P. Wallis, L.J. Wallis & R.D. Gray (2000). Seabird and louse coevolution: complex histories revealed by 12S rRNA sequences and reconciliation analyses. Systematic Biology 49 (3): 383—399
- ↑ Diptera: информация на сайте Красной книги МСОП (англ.)
Ссылки
- Britannica Online Encyclopedia: Lice (Phthiraptera)
- Discover Life — Insecta: Phthiraptera
- Phthiraptera in The Tree of Life Web Project
- Phthiraptera Central















