
Фотоси́нтез — сложный химический процесс преобразования энергии видимого света в энергию химических связей органических веществ при участии фотосинтетических пигментов.

Тилакоиды — ограниченные мембраной компартменты внутри хлоропластов и цианобактерий. В тилакоидах происходят светозависимые реакции фотосинтеза. Слово «тилакоид» происходит от греческого слова θύλακος, означающего «мешочек». Тилакоиды состоят из мембраны, окружающей просвет тилакоида. Тилакоиды хлоропластов часто имеют структуру, напоминающую стопку дисков. Эти стопки называют гранами. Граны соединены межграновыми или строматическими тилакоидами (ламеллами) в единое функциональное пространство.

Феофитин — химическое соединение, служащее одним из первых акцепторов электрона в цепи переноса электронов в реакционном центре фотосистемы II (ФСII) у растений, и в реакционном центре (РЦ) пурпурных бактерий. Как в реакционном центре ФСII, так и бактерий (P870), возбуждённые электроны передаются от воды на феофитин, который затем передаёт их хинону (QA). В целом механизмы, функции и задачи молекулы феофитина аналогичны друг другу в обеих транспортных цепях.

Хлорофи́лл a — особая форма хлорофилла, используемая для оксигенного фотосинтеза. Сильнее всего поглощает свет в фиолетово-голубой и оранжево-красной части спектра. Этот пигмент жизненно необходим для фотосинтеза в клетках эукариот, цианобактерий и прохлорофитов из-за своей способности отдавать возбуждённые электроны в электрон-транспортную цепь. Хлорофилл a также является частью антенного комплекса и передаёт резонансную энергию, которая затем поступает в реакционный центр, где расположены специальные хлорофиллы P680 и P700.

Светособирающие комплексы — пигмент-белковые комплексы фотосинтезирующих организмов, локализованные в фотосинтетических мембранах и выполняющие функцию первичного поглощения квантов света с последующей миграцией энергии возбуждения к реакционным центрам фотосистем. Также они обеспечивают тонкую настройку фотосинтетического аппарата и участвуют в его защите от фотоповреждений.

Фотосисте́ма I, или пластоциани́н-ферредокси́н-оксидоредукта́за — второй функциональный комплекс электрон-транспортной цепи (ЭТЦ) хлоропластов. Он принимает электрон от пластоцианина и, поглощая световую энергию, формирует сильный восстановитель П700, способный через цепь переносчиков электронов осуществить восстановление НАДФ+. Таким образом, при участии ФСI синтезируется источник электронов (НАДФН) для последующих реакций восстановления углерода в хлоропластах в цикле Кальвина. Кроме того, ФСI может осуществлять циклический транспорт электронов, сопряжённый с синтезом АТФ, обеспечивая дополнительный синтез АТФ в хлоропластах.

Пластоцианин — медьсодержащий белок, вовлечённый в транспорт электронов от фотосистемы II к фотосистеме I. Этот мономерный белок, состоящий у большинства сосудистых растений из 99 аминокислот, имеет молекулярную массу около 10,5 кДа. Он является представителем пластоцианинового семейства медьсвязывающих белков.

Реакционный центр — комплекс белков, пигментов и других кофакторов, взаимодействие которых обеспечивает реакцию превращения энергии света в химическую при фотосинтезе. Реакционный центр получает энергию или через непосредственное возбуждение одной из своих молекул или через перенос энергии от светособирающих комплексов, что даёт начало цепочке химических реакций, происходящей на связанных белками кофакторах. Эти кофакторы — светопоглощающие молекулы такие как хлорофилл, феофитин и хиноны. Энергия фотона используется для поднятия электрона на более высокий энергетический уровень. Запасённая таким образом свободная энергия идёт на восстановление цепочки акцепторов электрона с более высоким редокс-потенциалом.

Фотосисте́ма II (втора́я фотосисте́ма, фотосисте́ма два, ФСII), или H2O-пластохиноноксидоредуктаза — первый функциональный комплекс электрон-транспортной цепи (ЭТЦ) хлоропластов. Он расположен в мембранах тилакоидов всех растений, водорослей и цианобактерий. Поглощая энергию света в ходе первичных фотохимических реакций, он формирует сильный окислитель — димер хлорофилла a (П680+), который через цепь окислительно-восстановительных реакций способен вызвать окисление воды.

Цитохро́м-b6f-ко́мплекс, или пластохинолпластоцианинредукта́за — мультибелковый комплекс, который осуществляет окисление пластохинолов и восстановление белка пластоцианина, обеспечивая, таким образом, транспорт электронов между реакционными центрами фотосистемы I (ФСI) и фотосистемы II (ФСII). Он восстанавливает маленький водорастворимый белок пластоцианин, который переносит электрон к ФСII. Аналогичную реакцию катализирует цитохром-bc1-комплекс электрон-транспортной цепи митохондрий. Цитохром-b6f-комплекс присутствует в тилакоидной мембране хлоропластов растений, водорослей и цианобактерий. Он функционально объединяет две фотосистемы в единую цепь переноса электронов от воды к НАДФ+, то есть является участником нециклического потока электронов. Кроме того, цитохромный комплекс вовлечён в циклический транспорт электронов, осуществляемый фотосистемой I.
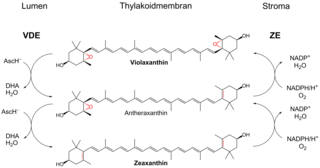
Ксантофилловый цикл, или виолаксантиновый цикл, в случае высших растений, выполняет функцию защиты фотосинтетического аппарата от избытка энергии при повышенной инсоляции. Он позволяет избежать фотоингибирования, значительно увеличив нефотохимическое тушение. Цикл включает в себя ферментативные взаимопревращения между виолаксантином и зеаксантином.
Нефотохимическое тушение — механизм защиты фотосинтетического аппарата от света высокой интенсивности, используемый растениями и водорослями. Суть процесса заключается в поглощении избыточной энергии (тушении) синглетного возбужденного хлорофилла молекулой-акцептором с последующим переходом этой молекулы в основное энергетическое состояние при помощи усиленной внутренней конверсии. Благодаря внутренней конверсии избыточная энергия возбуждения рассеивается в виде тепла, то есть расходуется на молекулярные колебания. Нефотохимическое тушение есть почти у всех фотосинтезирующих эукариот и цианобактерий. Оно помогает регулировать и защищать фотосинтетический аппарат в условиях, когда поглощается больше света, чем может быть непосредственно использовано в фотосинтезе.

Фотофосфорили́рование — процесс синтеза АТФ из АДФ за счёт энергии света. Как и в случае окислительного фосфорилирования, энергия света расходуется на создание протонного градиента на мембране тилакоидов или клеточной мембране бактерии, который затем используется АТФ-синтазой. Фотофосфорилирование — очень древняя форма фотосинтеза, которая есть у всех фототрофных эукариот, бактерий и архей. Различают два типа фосфорилирования — циклическое, сопряжённое с циклическим потоком электронов в электрон-транспортной цепи, и нециклическое, сопряжённое с прямым потоком электронов от H2O к НАДФ+ в случае эукариот или другого донора электрона в случае бактерий, например, H2S. Как разновидность нециклического типа выделяют псевдоциклическое фотофосфорилирование, при котором акцептором электронов служит кислород.

Фотосистема представляет собой функциональную и структурную единицу белковых комплексов, которые осуществляют первичные фотохимические реакции фотосинтеза: поглощение света, преобразование энергии и перенос электронов. Фотосистемы находятся в тилакоидной мембраны растений, водорослей и цианобактерий, либо в цитоплазматической мембране фотосинтезирующих бактерий. В общем случае все фотосистемы подразделяют на два типа — подобные фотосистеме II и подобные фотосистеме I.

Фотоингибирование — индуцируемое светом снижение фотосинтетической активности растений, водорослей или цианобактерии. Фотосистема II (ФСII) более чувствительна к свету, чем другие фотосинтетические машины, поэтому большинство исследователей определяют термин как светоиндуцируемое повреждения ФСII. В живых организмах, поврежденные избыточным освещением фотосистемы II постоянно репарируются за счёт деградации и синтеза белка D1 фотосинтетического реакционного центра ФСII. Понятие фотоингибирование также используется в более широком смысле, в значении «динамическое фотоингибирование», чтобы описать все реакции, которые приводят к снижению эффективности фотосинтеза, когда растения подвергаются воздействию света.
Эффект Каутского — явление изменения свечения хлорофилла, которое возникает при освещении предварительно адаптированного к темноте листа растения. Этот феномен был открыт Х. Каутским и А. Хиршем в 1931 году.

Флуоресценция хлорофилла — явление свечения хлорофилла при поглощении им света, происходит в результате возвращения молекулы из возбуждённого в основное состояние. Широко используется как показатель фотосинтетического преобразования энергии у высших растений, водорослей и бактерий. Возбуждённый хлорофилл теряет поглощённую световую энергию, растрачивая её на фотосинтез, переводя её в тепло в результате нефотохимического тушения или излучая в виде флуоресценции. Поскольку все эти процессы конкурируют друг с другом, анализируя флуоресценцию хлорофилла, можно получить представления об интенсивности фотосинтеза и здоровье растения.
П700 или первичный донор фотосистемы I — специальная пара хлорофиллов а в реакционном центре фотосистемы I с максимумом поглощения 700 нм. П700 представляет собой димер хлорофилла а и хлорофилла а′ у которого кетоэфирная группа в V кольце находится в цис-положении относительно плоскости молекулы. Наличие цис-кетоэфирной группы делает возможным образование димера из двух хлорофиллов посредством формирования водородных связей. П700 поглощает фотоны или получает энергию от антенных комплексов и использует её, чтобы поднять электроны на более высокий уровень. Далее электрон в ходе окислительно-восстановительной реакции переходит на цепь переносчиков. В окисленном состоянии окислительно-восстановительный редокс-потенциал П700 составляет +0,52 В, а в фотовозбуждённом состоянии он становится −1,2 В, то есть формируется мощный восстановитель, обеспечивающий восстановление НАДФ+.
Цитохром b559 — гетеродимерный белок, состоящий из одной альфа (PsbE) и одной бета (PsbF) субъединицы, между которыми расположен гем; важный компонент фотосистемы II.

Электрохими́ческий градие́нт, или градие́нт электрохимического потенциа́ла, — совокупность градиента концентрации и мембранного потенциала, которая определяет направление движения ионов через мембрану. Состоит из двух составляющих: химического градиента, или разницы в концентрациях растворённого вещества по обе стороны мембраны, и электрического градиента, или разницы зарядов, расположенных на противоположных сторонах мембраны. Градиент возникает вследствие неодинаковой концентрации ионов на противоположных сторонах водопроницаемой мембраны. Ионы двигаются через мембрану из области, имеющую более высокую концентрацию в область с более низкой концентрацией путём простой диффузии. Также ионы несут электрический заряд, который формирует электрический потенциал на мембране. Если существует неравномерное распределение зарядов по обе стороны мембраны, то разница в электрическом потенциале порождает силу, которая приводит к ионной диффузии, пока заряды по обе стороны не будут сбалансированы.
