Рапетозавр
| † Рапетозавр | |||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 Скелет молодой особи в музее естественной истории им. Филда | |||||||||||||||||||||||||||||||||||||||||
| Научная классификация | |||||||||||||||||||||||||||||||||||||||||
Домен: Эукариоты Царство: Животные Подцарство: Эуметазои Без ранга: Двусторонне-симметричные Без ранга: Вторичноротые Тип: Хордовые Подтип: Позвоночные Инфратип: Челюстноротые Надкласс: Четвероногие Клада: Амниоты Клада: Завропсиды Клада: Архозавры Клада: Авеметатарзалии Клада: Динозавроморфы Клада: Динозавры Клада: Ящеротазовые Подотряд: † Завроподоморфы Клада: Клада: Инфраотряд: † Завроподы Клада: † Макронарии Клада: † Титанозавриформы Клада: Клада: † Титанозавры Клада: † Литостротии Род: † Рапетозавр | |||||||||||||||||||||||||||||||||||||||||
| Международное научное название | |||||||||||||||||||||||||||||||||||||||||
| Rapetosaurus Curry Rogers & Forster, 2001 | |||||||||||||||||||||||||||||||||||||||||
| Единственный вид | |||||||||||||||||||||||||||||||||||||||||
Curry Rogers & Forster, 2001 | |||||||||||||||||||||||||||||||||||||||||
| Геохронология Маастрихтский век 72,1—66,0 млн лет
◄ Наше время◄ Мел-палеогеновое вымирание ◄ Триасовое вымирание◄ Массовоепермское вымирание ◄ Девонское вымирание◄ Ордовикско-силурийское вымирание◄ Кембрийский взрыв | |||||||||||||||||||||||||||||||||||||||||
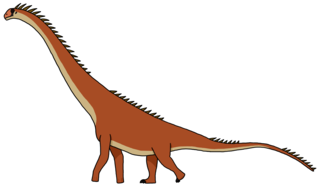
Рапетозавр[1] (лат. Rapetosaurus) — род завропод из клады Lithostrotia, живших во времена позднемеловой эпохи (маастрихтский век) на территории Мадагаскара[2]. Известен один вид — Rapetosaurus krausei.
Характеристики

Рапетозавр был типичным завроподом: имел длинную шею, тонкий хвост и крупное слоноподобное тело. Был довольно скромного размера для титанозавров: молодые особи достигали 8 м в длину и весили около 5—6 тонн, взрослые животные были в два раза крупнее — 15[3] м длиной и 30—50 тонн массой, что по размеру вполовину меньше таких титанозавров, как аргентинозавр или паралититан.
Череп имел сходство с черепами диплодоцид — удлинённый и узкий, ноздри были расположены на вершине черепа. Важной отличительной особенностью в строении черепа рапетозавра являлись расширенные анторбитальные окна (отверстия перед глазницами), которые доходили до уровня начала альвеолярной дуги. Челюсти содержали маленькие карандашоподобные зубы, пригодные для срывания листьев с деревьев.
Позвонки завропод являются наиболее важной частью скелета, так как содержали множественные полости с воздушными мешками определённой формы для каждого рода, позволяющим палеонтологам определить филогенетические связи животного. В шейном отделе позвоночника рапетозавра содержалось 15 позвонков, причём первые 5 из них были удлинённые, а последующие становились более короткими и широкими. Эти 10 шейных позвонков были опистоцельными — тела позвонков выпуклы спереди и вогнуты сзади. Также опистоцельными были и 6 тазовых позвонков, причём сращения между ними отсутствовали. Общее количество хвостовых позвонков неизвестно, найдено было только 18 штук. Хвостовые позвонки были процельными (их тела вогнуты спереди и выпуклы сзади) с длинными остистыми отростками, направленными вверх.
Таз рапетозавра характеризуется короткой седалищной костью — 54 % от длины лобковой кости. Вертлужная впадина тазобедренного сустава была образована в большой степени подвздошной и лобковой костями, и лишь небольшим фрагментом седалищной кости. Кости стопы анатомически соединены, самым длинным пальцем из пяти являлся третий. Пятый палец был треугольной формы и значительно уменьшен.
Открытие и описание

Ископаемые остатки были найдены недалеко от портового города Махадзанга на северо-западе Мадагаскара, в слоях песчаника, известные как пачка Анембалемба (часть формации Маеварано), датированная маастрихтским веком позднемеловой эпохи (возрастом около 70 миллионов лет). Окаменелости нашла полевая группа из Университета штата Нью-Йорк в Стоуни-Брук совместно с Университетом Антананариву. Руководитель группы Дэвид Крауз занимался раскопками с 1993 года.
Открытие рапетозавра стало первым случаем обнаружения титанозавра с почти совершенно неповреждённым скелетом и полностью сохранившемся черепом. Данная находка смогла прояснить многие проблемы классификации этой группы завропод, а также стала основой для реконструкции многих других титанозавров, в основном известных только по разрозненным ископаемым остаткам.
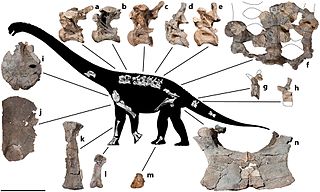
В результате раскопок были обнаружены два частично сохранившихся черепа взрослых особей (голотип UA 8698 состоял из правой половины верхней челюсти с 8 зубами и левой половиной нижней челюсти с 11 зубами), скелет молодой особи (голотип FMNH PR2209), у которого не хватало только несколько хвостовых позвонков. Именно голотип FMNH PR2209 является самыми хорошо сохранившемся ископаемыми остатками титанозавров, из когда либо обнаруженных.
Описание было опубликовано в 2001 году Кристиной Карри Роджерс и Катериной А. Форстер в Nature по практически полному скелету молодой особи и частичным ископаемым остаткам трёх других экземпляров. Родовое название дано в честь гигантского божества Рапето из малагасийского фольклора, а видовой эпитет в честь Дэвида Крауза — руководителя полевой группы.
Классификация
Рапетозавр был типичным представителем титанозавров (Titanosauria) — группы, к которой принадлежали одни из последних живших завропод и самые крупные животные, когда либо существовавшие. При описании Карри Роджерс и Катерина Фостер отметили сходства черепа рапетозавра с черепами немегтозавра и кваэзитозавра из позднего мела Монголии[4]. На основании этого Уилсон в 2002 году отнёс рапетозавра к семейству нэмэгтозаврид[5]. В 2003 году Уилсон вместе с Апчёрчем предложил признать семейство титанозаврид недействительным на основании того, что род Titanosaurus был признан nomen dubium, так как голотип не соответствовал диагностическим характеристикам из-за плохого состояния находки и малого количества ископаемого материала[6]. Вместо Titanosauridae Апчёрч в 2004 году создал новую кладу Lithostrotia, куда был помещён в том числе и рапетозавр[7]. В 2005 году Карри Роджерс публикует статью о филогении титанозавров, в которой кладу Lithostrotia относит к семейству сальтозаврид[8]. В том же году Уилсон повторно помещает рапетозавра как род в составе нэмэгтозаврид[9]. В 2011 году бразильские палеонтологи во главе с Заэром включили нэмэгтозаврид в состав клады Lithostrotia[10]. По исследованиям 2016 года[11], считается, что рапетозавр является паралифетическим родом по отношению к нэмэгтозавридам и поэтому является incertae sedis в составе Lithostrotia.
Палеобиология
Рост

Кристина Роджерс и её коллеги в 2016 году обнаружили в музейной коллекции новые образцы совсем юных особей рапетозавров[12]. По оценкам особь весила около 40 кг и к моменту смерти, вероятно, имела возраст от 39 до 77 дней и умерла вследствие засухи от голода. По подсчётам во время вылупления из яйца детёныш весил 3,4 кг. Изучение микроструктуры костей показало, что у детёнышей этих завропод был очень быстрый рост, они были самостоятельны и не нуждались в родительской заботе.
Палеоэкология

Во время маастрихтского века Мадагаскар, как и в нынешнее время, был островом, отделившемся от Индийского субконтинента за 20 миллионов лет до этого. Он дрейфовал на север, однако находился всё ещё на 10-15о южнее, чем в наши дни. Климат тогда был полузасушливым, с выраженной сезонностью температур и количества осадков. Многие животные обитали в прибрежной пойме, изрезанной большим количеством песчаных речных русел[13]. Геологические данные говорят о том, что по этим руслам в сезон дождей возникали сильные водные потоки, уносящие с собой ископаемые остатки организмов, погибших в предшествующий сухой сезон. Такие условия обеспечивали исключительную сохранность окаменелостей[14]. Уровень моря в этом районе постепенно повышался в течение всего маастрихта и палеоцена, так что, вероятно, рапетозавры бродили в зонах прилива. Соседняя формация Беривотра представляет уже из себя морские отложения[13].
Помимо рапетозавра, в формации Маеварано были обнаружены рыбы, лягушки, ящерицы, змеи[13], семь видов крокодиломорфов[15], Vorona[16] и несколько других птиц[13], летающий дромеозаврид рахонавис[17][18], ноазаврид масиакозавр[19] и абелизаврид майюнгозавр[20]. Также было обнаружено большое количество млекопитающих, такие как неплацентарные эутерии и гондванатерии, некоторые из которых, например винтана, достигали довольно крупных размеров.

Череп майюнгазавра был обнаружен в 1996 году. Этот абелизаврид похож на других представителей семейства, найденных в Индии и Аргентине, что указывает на существование сухопутных мостов между фрагментами бывшего суперконтинента Гондвана в конце мелового периода, гораздо позже, чем считалось ранее. Вероятнее всего, существовал перешеек, который позволял животным перемещаться из Южной Америки и Антарктиду, а в дальнейшем в Индию и Мадагаскар. Майюнгозавр был крупнейшим хищником на данной территории, в то время как единственные известные крупные растительноядные животные были завроподы, подобные рапетозавру. Учёные предполагают, что майюнгозавры специализировались в охоте на завропод. Следы зубов майюнгозавров на костях рапетозавров указывают на то, что они питались этими завроподами[21].
Как правило, титанозавры были необычны среди завропод тем, что могли сосуществовать вместе с крупными птицетазовыми, такими как цератопсиды, гадрозавры и анкилозавры. Тем не менее, рапетозавр был нетипичным среди титанозавров в том, что он разделял бассейн Махадзанга с другим крупным завроподом — Vahiny depereti[22]. Более мелкие растительноядные животные были редки — только один, симозух, обнаружен за более чем вековые раскопки в этом районе. Из-за отсутствия птицетазовых, предполагается, что меловой Мадагаскар имел совсем другие растительноядные сообщества животных, чем в других частях планеты[4].
Примечания
- ↑ Татаринов Л. П. Очерки по эволюции рептилий. Архозавры и зверообразные. — М. : ГЕОС, 2009. — С. 124. — 377 с. : ил. — (Труды ПИН РАН ; т. 291). — 600 экз. — ISBN 978-5-89118-461-9.
- ↑ Rapetosaurus (англ.) информация на сайте Paleobiology Database. (Дата обращения: 12 октября 2017).
- ↑ Montague R. Estimates of body size and geological time of origin for 612 dinosaur genera (Saurischia, Ornithischia) (англ.) // Florida Scientist : journal. — 2006. — Vol. 69, no. 4. — P. 243—257.
- ↑ 1 2 Catherine A. Forster, Kristina Curry Rogers. The last of the dinosaur titans: a new sauropod from Madagascar (англ.) // Nature. — 2001-08. — Vol. 412, iss. 6846. — P. 530—534. — ISSN 1476-4687. — doi:10.1038/35087566. Архивировано 12 апреля 2019 года.
- ↑ J. A. Wilson. Sauropod dinosaur phylogeny: critique and cladistic analysis. (англ.) // Zoological Journal of the Linnean Society[англ.]. — Oxford University Press, 2002. — No. 136. — P. 217—276. Архивировано 9 февраля 2022 года.
- ↑ J. A. Wilson and P. Upchurch. A revision of Titanosaurus Lydekker (Dinosauria – Sauropoda), the first dinosaur genus with a 'Gondwanan' distribution. (англ.) // Journal of Systematic Palaeontology[англ.]. — Taylor & Francis. — No. 1 (3). — P. 125—160. Архивировано 15 февраля 2019 года.
- ↑ P. Upchurch, P. M. Barrett, and P. Dodson. The Dinosauria (2nd edition).. — University of California Press, Berkeley, 2004. — С. 259—322.
- ↑ K. A. Curry Rogers, J. A. Wilson. The Sauropods: Evolution and Paleobiology.. — University of California Press, Berkeley, 2005. — С. 50—103.
- ↑ (PDF) Redescription of the Mongolian sauropod Nemegtosaurus mongoliensis Nowinski (Dinosauria: Saurischia) and comments on Late Cretaceous sauropod diversity (англ.). ResearchGate. Дата обращения: 14 февраля 2019. Архивировано 29 октября 2021 года.
- ↑ Hussam Zaher, Diego Pol, Alberto B. Carvalho, Paulo M. Nascimento, Claudio Riccomini. A complete skull of an early cretaceous sauropod and the evolution of advanced titanosaurians // PloS One. — 2011-02-07. — Т. 6, вып. 2. — С. e16663. — ISSN 1932-6203. — doi:10.1371/journal.pone.0016663. Архивировано 15 февраля 2019 года.
- ↑ Juan P. Coria, Jorge O. Calvo, Leonardo D. Ortiz David, Matthew C. Lamanna, Bernardo J. González Riga. A gigantic new dinosaur from Argentina and the evolution of the sauropod hind foot (англ.) // Scientific Reports. — 2016-01-18. — Vol. 6. — P. 19165. — ISSN 2045-2322. — doi:10.1038/srep19165. Архивировано 30 января 2019 года.
- ↑ (PDF) Precocity in a tiny titanosaur from the Cretaceous of Madagascar (англ.). ResearchGate. Дата обращения: 13 февраля 2019. Архивировано 14 февраля 2019 года.
- ↑ 1 2 3 4 Rogers, Raymond R.; Krause, David W.; Curry Rogers, Kristina; Rasoamiaramanana, Armand H.; Rahantarisoa, Lydia. PALEOENVIRONMENT AND PALEOECOLOGY OF MAJUNGASAURUS CRENATISSIMUS (THEROPODA: ABELISAURIDAE) FROM THE LATE CRETACEOUS OF MADAGASCAR. Journal of Vertebrate Paleontology. www.tandfonline.com (2007). Дата обращения: 13 февраля 2019.
- ↑ Raymond R. Rogers. Fine-grained debris flows and extraordinary vertebrate burials in the Late Cretaceous of Madagascar (англ.) // Geology. — 2005-04-01. — Vol. 33, iss. 4. — P. 297—300. — ISSN 0091-7613. — doi:10.1130/G21036.1. Архивировано 14 февраля 2019 года.
- ↑ Raymond R. Rogers, Gregory A. Buckley, Scott D. Sampson, Kristina Curry Rogers, Patrick M. O'Connor. [178:LCTVFM2.0.CO;2/LATE-CRETACEOUS-TERRESTRIAL-VERTEBRATES-FROM-MADAGASCAR--IMPLICATIONS-FOR-LATIN/10.3417/0026-6493(2006)93[178:LCTVFM]2.0.CO;2.full LATE CRETACEOUS TERRESTRIAL VERTEBRATES FROM MADAGASCAR: IMPLICATIONS FOR LATIN AMERICAN BIOGEOGRAPHY1] // Annals of the Missouri Botanical Garden. — 2006/08. — Т. 93, вып. 2. — С. 178—208. — ISSN 2162-4372 0026-6493, 2162-4372. — doi:10.3417/0026-6493(2006)93[178:LCTVFM]2.0.CO;2.
- ↑ Scott D. Sampson, David W. Krause, Luis M. Chiappe, Catherine A. Forster. The first Cretaceous bird from Madagascar (англ.) // Nature. — 1996-08. — Vol. 382, iss. 6591. — P. 532—534. — ISSN 1476-4687. — doi:10.1038/382532a0. Архивировано 3 сентября 2020 года.
- ↑ null Forster, null Sampson, null Chiappe, null Krause. The theropod ancestry of birds: new evidence from the late cretaceous of madagascar // Science (New York, N.Y.). — 1998-03-20. — Т. 279, вып. 5358. — С. 1915—1919. — ISSN 1095-9203. Архивировано 14 февраля 2019 года.
- ↑ Federico L. Agnolín, Sebastián Apesteguía, Peter J. Makovicky. The earliest dromaeosaurid theropod from South America (англ.) // Nature. — 2005-10. — Vol. 437, iss. 7061. — P. 1007—1011. — ISSN 1476-4687. — doi:10.1038/nature03996. Архивировано 27 апреля 2019 года.
- ↑ Catherine A. Forster, Matthew T. Carrano, Scott D. Sampson. A bizarre predatory dinosaur from the Late Cretaceous of Madagascar (англ.) // Nature. — 2001-01. — Vol. 409, iss. 6819. — P. 504—506. — ISSN 1476-4687. — doi:10.1038/35054046. Архивировано 24 октября 2019 года.
- ↑ Krause, David W.; Sampson, Scott D.; Carrano, Matthew T.; O'Connor, Patrick M. Overview of the history of discovery, taxonomy, phylogeny, and biogeography of Majungasaurus crenatissimus (Theropoda: Abelisauridae) from the Late Cretaceous of Madagascar. Journal of Vertebrate Paleontology. www.tandfonline.com (2007). Дата обращения: 13 февраля 2019.
- ↑ Kristina Curry Rogers, David W. Krause, Raymond R. Rogers. Cannibalism in the Madagascan dinosaur Majungatholus atopus (англ.) // Nature. — 2003-04. — Vol. 422, iss. 6931. — P. 515—518. — ISSN 1476-4687. — doi:10.1038/nature01532. Архивировано 6 ноября 2021 года.
- ↑ Rogers, K. C.; Wilson, J. A. Vahiny depereti, gen. et sp. nov., a new titanosaur (Dinosauria, Sauropoda) from the Upper Cretaceous Maevarano Formation, Madagascar. Journal of Vertebrate Paleontology. www.tandfonline.com (2014). Дата обращения: 16 февраля 2019.