Рецептор эритропоэтина
| Рецептор эритропоэтина | |||
|---|---|---|---|
 | |||
| Доступные структуры | |||
| PDB | Поиск ортологов: PDBe, RCSB | ||
| Идентификаторы | |||
| Символ | EpoR, Epo-R | ||
| Внешние ID | OMIM: 133171 MGI: 95408 HomoloGene: 1731 ChEMBL: 1817 GeneCards: Ген EpoR, Epo-R | ||
| Профиль экспрессии РНК | |||
 | |||
 | |||
 | |||
| Больше информации | |||
| Ортологи | |||
| Вид | Человек | Мышь | |
| Entrez | 2057 | 13857 | |
| Ensembl | ENSG00000187266 | ENSMUSG00000006235 | |
| UniProt | P19235 | P14753 | |
| RefSeq (мРНК) | NM_000121 | NM_010149 | |
| RefSeq (белок) | NP_000112 | NP_034279 | |
| Локус (UCSC) | Chr 19: 0 – 0 Mb | Chr 9: 0 – 0 Mb | |
| Поиск в PubMed | Искать | Искать | |
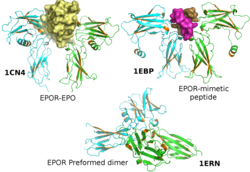
Рецептор эритропоэтина (EpoR) — белок, который кодируется у людей геном EpoR[1]. Пептидная цепь EpoR имеет атомную массу 52kDa, атомная масса гликопептида вместе с единственной углеводной цепью - 56-57kDa (по другим данным [1], 66-105kDa). EpoR принадлежит к семейству цитокиновых рецепторов. EpoR присутствует на мембране в виде гомодимеров[2], которые при связывании с лигандом эритропоэтином (Epo) меняют свою конформацию. Эти изменения конформации вызывают аутофосфорилирование киназы Jak2, которая связана с ним изначально, это связано с активностью Jak2[3][4]. Сегодня наиболее аргументированной функцией EpoR является способствование распространению эритроидных предшественников и спасению их от апоптоза[1]. Субъединицы EpoR также способны образовывать гетеродимеры с другими белками-рецепторами — βcR и EPHB4.
Механизм действия

Цитоплазматические домены EpoR содержат ряд фосфотирозинов, фосфорилированных Jak2 и служащих местами стыковки для различных активаторов внутриклеточного пути.
В дополнение к активации Ras/Akt и ERK/MAP киназы, фосфатидилинозитол 3-киназы/AKT пути и STAT транскрипционных факторов, фосфотирозины также служат в качестве стыковочных участков для фосфатаз, отрицательно воздействующих на сигнализацию EpoR, что предотвращает сверхактивацию.
Выживание клеток эритроидного ряда
Главная роль EpoR — стимулировать быстрое распространение клеток-предшественников эритроцитов и спасение этих клеток от гибели[5].
EpoR вместе с фактором транскрипции GATA-1, вызывает транскрипцию белков, способствующих сохранению Bcl-xL[6].
Кроме того, EpoR задействован в подавлении экспрессии рецепторов гибели Fas, Trail и TNFa, которые негативно влияют на эритропоэз[7][8][9].
До сих пор неизвестно, непосредственно ли Epo/EpoR служит причиной распространения и дифференцировки предшественников эритроцитов в естественных условиях, поскольку эффекты были описаны на основе работы in vitro[5].
Дифференцировка эритроидного ряда
Есть основания полагать, что дифференцировка эритроидного ряда в основном зависит от присутствия и индукции таких транскрипционных факторов, как GATA-1, FOG-1 и EKLF, а также от супрессии таких миелоидных и лимфоидных факторов, как PU.1.[10] Прямые эффекты сигнализации EpoR — индукция эритроид-специфичных генов, как бета-глобин, в основном плохо поддаются изучению. Известно, что GATA-1 может провоцировать экспрессию EpoR.[11] В свою очередь, сигнальный путь PI3-K/AKT увеличивает активность GATA-1.[12]
Клеточный цикл эритроидного ряда
Распространение EpoR, вероятнее всего, зависит от типа клеток. Известно, что EpoR может активировать митогенные сигнальные пути и управлять распространением разнообразных неэритроидных и раковых клеток.
Благодаря сигнализации EpoR, CFU-e-предшественники входят в клеточный цикл во время индукции GATA-1 и подавления PU.1.[13] Во время следующих этапов дифференцировки размер клетки уменьшается, а в самом конце ядро выбрасывается наружу. Выживание клеток на этих этапах как раз и зависит от сигнализации EpoR. Также сигнализация EpoR влияет на распространения BFU-e предшественников, которые еще не были достаточно хорошо изучены.
Кроме того, из некоторых данных о макроцитозе при гипоксическом стрессе (когда Epo увеличивается в тысячи раз) следует, что в последующих этапах митоза практически нет, а экспрессия EpoR очень низкая (или отсутствует). Это нужно для того, чтобы как можно скорее обеспечить доступ к запасу эритроцитов. Эти данные доказывают, что ограниченная способность распространяться зависит от Epo, а не от других факторов. EpoR в дифференцировке эритроидного ряда может функционировать в первую очередь как фактор выживания, в то время как его влияние на клеточный цикл в естественных условиях проявляется по прошествии некоторого времени.[14] В других клеточных системах EpoR может обеспечить определенный пролиферативный сигнал. [15]
Участие мультипотентных предшественников в дифференцировке эритроидного ряда
В данное время роль EpoR в дифференцировке неясна. Экспрессия EpoR может увеличиваться ещё в отделе гемопоэтических стволовых клеток[16]. Неизвестно, какую роль играет сигнализация EpoR в ранней стадии производства эритробластов: разрешающую (то есть индуцирующую только выживание) или инструктирующую (то есть активирующую маркёров для блокировки предшественники на заданной траектории дифференцировки).
Текущие публикации предполагают, что в первую очередь они играют разрешающую роль. Производство BFU-e и CFU-e предшественников было нормальным в эмбрионах грызунов с нокаутированным Epo, равно как и с нокаутированным EpoR[17]. Однако при добавлении Epo или при гипоксическом стрессе число BFU-e и CFU-e крайне сильно возрастает. В любом случае, неясно, какую из двух ролей всё же играет EpoR. Дополнительные вопросы вызывает информация о том, что пути, которые активирует EpoR, общие со многими другими рецепторами. А если заменить EpoR на пролактиновый рецептор, то всё равно идет поддержка дифференцировки и выживания эритроидного ряда, но эти данные, опять же, получены из исследований in vitro[18][19]. В итоге эти данные говорят о том, что, вероятнее всего EpoR участвует в дифференцировке эритроидного ряда не неизвестной инструктирующей функцией, а его ролью в выживании мультипотентных предшественников.
Исследование мутаций на животных
Мыши с усеченным EpoR жизнеспособны[20], что дает возможность предположить, что активность Jak2 достаточна для обеспечения эритропоэза без обязательного фосфотирозинового молекулярного докинга.
Мыши с вариантом рецептора EpoR-HM обладают мутировавшем из тирозина в 343 позиции фенилаланином, что делает молекулярный докинг Stat5 неэффективным. Эти мыши страдают анемией и дают слабый ответ на гипоксический стресс.
Мыши с нокаутированным EpoR имеют дефекты в сердце, мозге и сосудистой системе.
Клиническое значение
Повышенное производство эритроцитов повышает шанс развития таких патологий, как тромбоз и апоплексический удар. Дефекты EpoR могут приводить к эритролейкозу и наследственному эритроцитозу. Мутации в Jak2-киназах, связанные с EpoR, также могут привести к истинной полицитемии.[21]
Редко подобное сверхпроизводство эритроцитов просто повышает выносливость без отрицательных эффектов.[22]
Источники
- ↑ 1 2 Entrez Gene: EPOR erythropoietin receptor.
- ↑ Livnah O., Stura E.A., Middleton S.A., Johnson D.L., Jolliffe L.K., Wilson I.A. Crystallographic evidence for preformed dimers of erythropoietin receptor before ligand activation (англ.) // Science : journal. — 1999. — February (vol. 283, no. 5404). — P. 987—990. — doi:10.1126/science.283.5404.987. — PMID 9974392.
- ↑ Youssoufian H., Longmore G., Neumann D., Yoshimura A., Lodish H.F. Structure, function, and activation of the erythropoietin receptor (англ.) // Blood : journal. — American Society of Hematology[англ.], 1993. — May (vol. 81, no. 9). — P. 2223—2236. — PMID 8481505.
- ↑ Wilson I.A., Jolliffe L.K. The structure, organization, activation and plasticity of the erythropoietin receptor (англ.) // Current Opinion in Structural Biology : journal. — 1999. — December (vol. 9, no. 6). — P. 696—704. — doi:10.1016/S0959-440X(99)00032-9. — PMID 10607675.
- ↑ 1 2 Koury M.J., Bondurant M.C. Erythropoietin retards DNA breakdown and prevents programmed death in erythroid progenitor cells (англ.) // Science : journal. — 1990. — April (vol. 248, no. 4953). — P. 378—381. — doi:10.1126/science.2326648. — PMID 2326648.
- ↑ Socolovsky M., Fallon A.E., Wang S., Brugnara C., Lodish H.F. Fetal anemia and apoptosis of red cell progenitors in Stat5a-/-5b-/- mice: a direct role for Stat5 in Bcl-X(L) induction (англ.) // Cell : journal. — Cell Press, 1999. — July (vol. 98, no. 2). — P. 181—191. — doi:10.1016/S0092-8674(00)81013-2. — PMID 10428030.
- ↑ De Maria R., Testa U., Luchetti L., Zeuner A., Stassi G., Pelosi E., Riccioni R., Felli N., Samoggia P., Peschle C. Apoptotic role of Fas/Fas ligand system in the regulation of erythropoiesis (англ.) // Blood : journal. — American Society of Hematology[англ.], 1999. — February (vol. 93, no. 3). — P. 796—803. — PMID 9920828.
- ↑ Liu Y., Pop R., Sadegh C., Brugnara C., Haase V.H., Socolovsky M. Suppression of Fas-FasL coexpression by erythropoietin mediates erythroblast expansion during the erythropoietic stress response in vivo (англ.) // Blood : journal. — American Society of Hematology[англ.], 2006. — July (vol. 108, no. 1). — P. 123—133. — doi:10.1182/blood-2005-11-4458. — PMID 16527892. — PMC 1895827.
- ↑ Felli N., Pedini F., Zeuner A., Petrucci E., Testa U., Conticello C., Biffoni M., Di Cataldo A., Winkles J.A., Peschle C., De Maria R. Multiple members of the TNF superfamily contribute to IFN-gamma-mediated inhibition of erythropoiesis (англ.) // Journal of Immunology[англ.] : journal. — 2005. — August (vol. 175, no. 3). — P. 1464—1472. — doi:10.4049/jimmunol.175.3.1464. — PMID 16034083.
- ↑ Cantor A.B., Orkin S.H. Transcriptional regulation of erythropoiesis: an affair involving multiple partners (англ.) // Oncogene : journal. — 2002. — May (vol. 21, no. 21). — P. 3368—3376. — doi:10.1038/sj.onc.1205326. — PMID 12032775.
- ↑ Zon L.I., Youssoufian H., Mather C., Lodish H.F., Orkin S.H. Activation of the erythropoietin receptor promoter by transcription factor GATA-1 (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 1991. — December (vol. 88, no. 23). — P. 10638—10641. — doi:10.1073/pnas.88.23.10638. — PMID 1660143. — PMC 52985.
- ↑ Zhao W., Kitidis C., Fleming M.D., Lodish H.F., Ghaffari S. Erythropoietin stimulates phosphorylation and activation of GATA-1 via the PI3-kinase/AKT signaling pathway (англ.) // Blood : journal. — American Society of Hematology[англ.], 2006. — February (vol. 107, no. 3). — P. 907—915. — doi:10.1182/blood-2005-06-2516. — PMID 16204311. — PMC 1895894.
- ↑ Pop R., Shearstone J.R., Shen Q., Liu Y., Hallstrom K., Koulnis M., Gribnau J., Socolovsky M. A key commitment step in erythropoiesis is synchronized with the cell cycle clock through mutual inhibition between PU.1 and S-phase progression (англ.) // PLoS Biology : journal. — 2010. — Vol. 8, no. 9. — doi:10.1371/journal.pbio.1000484. — PMID 20877475. — PMC 2943437.
- ↑ Seno S., Miyahara M., Asakura H., Ochi O., Matsuoka K., Toyama T. MACROCYTOSIS RESULTING FROM EARLY DENUCLEATION OF ERYTHROID PRECURSORS (нем.) // Blood : magazin. — American Society of Hematology[англ.], 1964. — November (Bd. 24). — S. 582—593. — PMID 14236733.
- ↑ Borsook H., Lingrel J.B., Scaro J.L., Millette R.L. Synthesis of haemoglobin in relation to the maturation of erythroid cells (англ.) // Nature : journal. — 1962. — October (vol. 196, no. 4852). — P. 347—350. — doi:10.1038/196347a0. — PMID 14014098.
- ↑ Forsberg E.C., Serwold T., Kogan S., Weissman I.L., Passegué E. New evidence supporting megakaryocyte-erythrocyte potential of flk2/flt3+ multipotent hematopoietic progenitors (англ.) // Cell : journal. — Cell Press, 2006. — July (vol. 126, no. 2). — P. 415—426. — doi:10.1016/j.cell.2006.06.037. — PMID 16873070.
- ↑ Wu H., Liu X., Jaenisch R, Lodish H.F. Generation of committed erythroid BFU-E and CFU-E progenitors does not require erythropoietin or the erythropoietin receptor (англ.) // Cell : journal. — Cell Press, 1995. — October (vol. 83, no. 1). — P. 59—67. — doi:10.1016/0092-8674(95)90234-1. — PMID 7553874.
- ↑ Socolovsky M., Fallon A.E., Lodish H.F. The prolactin receptor rescues EpoR-/- erythroid progenitors and replaces EpoR in a synergistic interaction with c-kit (англ.) // Blood : journal. — American Society of Hematology[англ.], 1998. — September (vol. 92, no. 5). — P. 1491—1496. — PMID 9716574.
- ↑ Socolovsky M., Dusanter-Fourt I., Lodish H.F. The prolactin receptor and severely truncated erythropoietin receptors support differentiation of erythroid progenitors (англ.) // The Journal of Biological Chemistry : journal. — 1997. — May (vol. 272, no. 22). — P. 14009—14012. — doi:10.1074/jbc.272.22.14009. — PMID 9162017.
- ↑ Zang H., Sato K., Nakajima H., McKay C., Ney P.A., Ihle J.N. The distal region and receptor tyrosines of the Epo receptor are non-essential for in vivo erythropoiesis (англ.) // The EMBO Journal : journal. — 2001. — June (vol. 20, no. 12). — P. 3156—3166. — doi:10.1093/emboj/20.12.3156. — PMID 11406592. — PMC 150206.
- ↑ James C., Ugo V., Le Couédic J.P., Staerk J., Delhommeau F., Lacout C., Garçon L., Raslova H., Berger R., Bennaceur-Griscelli A., Villeval J.L., Constantinescu S.N., Casadevall N., Vainchenker W. A unique clonal JAK2 mutation leading to constitutive signalling causes polycythaemia vera (англ.) // Nature : journal. — 2005. — April (vol. 434, no. 7037). — P. 1144—1148. — doi:10.1038/nature03546. — PMID 15793561.
- ↑ de la Chapelle A., Träskelin A.L., Juvonen E. Truncated erythropoietin receptor causes dominantly inherited benign human erythrocytosis (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 1993. — May (vol. 90, no. 10). — P. 4495—4499. — doi:10.1073/pnas.90.10.4495. — PMID 8506290. — PMC 46538.