
Ген — в классической генетике — наследственный фактор, который несёт информацию об определённом признаке или функции организма, и который является структурной и функциональной единицей наследственности. В таком качестве термин «ген» был введён в 1909 году датским ботаником, физиологом растений и генетиком Вильгельмом Йоханнсеном.

Ми́кроРНК — малые некодирующие молекулы РНК длиной 18—25 нуклеотидов, обнаруженные у растений, животных и некоторых вирусов, принимающие участие в транскрипционной и посттранскрипционной регуляции экспрессии генов путём РНК-интерференции. Помимо внутриклеточной обнаружена внеклеточная (циркулирующая) микроРНК.
Бисульфи́тное секвени́рование — общее название группы методов, направленных на изучение паттерна метилирования ДНК посредством обработки её бисульфитом.

Ku80 — белок, закодированный у человека геном XRCC5. Вместе с белком Ku70 белок Ku80 образует гетеродимер, связывающийся с концами двунитевых разрывов в ДНК, и требуется для репарации ДНК путём негомологичного соединения концов (NHEJ). Также белок Ku требуется для V(D)J рекомбинации, которая использует путь NHEJ для создания разнообразия антиген-связывающих центров антител в иммунной системе млекопитающих.

G-квадру́плексы (англ. G-quadruplex, а также G-tetrads или G4) — последовательности нуклеиновых кислот, обогащенные гуанином и способные образовывать структуры из четырёх цепей. Цепи нуклеиновых кислот из гуанозиновых олиго- и полинуклеотидов способны связываться друг с другом при наличии моновалентного катиона небольшого размера, чаще всего — калия. С помощью дифракционного анализа было показано, что такие поли(G)-нити представляют собой новый тип укладки ДНК, четырёхцепочечную спираль, где четыре гуаниновых основания из разных цепей образуют плоскую структуру, удерживаемую парными взаимодействиями G-G (рис. 1). Такие структуры отличаются высокой стабильностью в растворе и называются гуаниновыми (G)-квартетами, или G-тетрадами. Каждый G-квартет скреплен в сумме восемью водородными связями, образованными взаимодействием Уотсон-Криковской стороны одного гуанинового основания с Хугстиновской стороной другого. G-квадруплексы могут быть также образованы короткими олигонуклеотидами с соответствующей последовательностью, которую можно схематически записать как GmXnGmXoGmXpGm, где m — количество гуанинов в G-блоке. Эти гуанины обычно непосредственно задействованы в образовании G-тетрад. Xn, Xo и Xp могут быть комбинацией любых остатков, включая G; такие участки формируют петли между G-тетрадами.
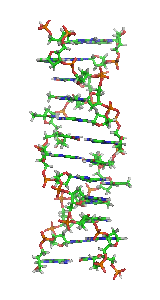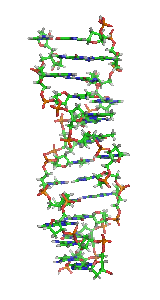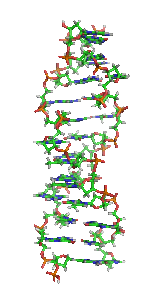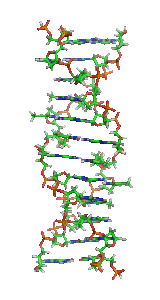
Z-ДНК — одна из многих возможных структур двойной спирали ДНК, представляет собой левозакрученную двойную спираль. Z-ДНК является одной из трёх биологически активных двойных спиральных структур ДНК, наряду с А-ДНК и В-ДНК, хотя точные её функции к настоящему моменту не определены.

Внутренняя терминация, также ρ-независимая терминация, ро-независимая терминация , — механизм остановки транскрипции гена у прокариот. Суть этого механизма такова: мРНК содержит гуанин-цитозин-обогащённую последовательность, которая может образовывать структуры типа шпилька в 7—20 пар оснований в длину. Гуанин и цитозин образуют друг с другом три водородные связи и поэтому связаны довольно прочно. Сразу после шпильки располагается участок, обогащённый урацилом. Связи между урацилом и аденином очень слабы. Белок, связанный с РНК-полимеразой (nusA), настолько прочно связывается со шпилькой, что это вызывает временную остановку полимеразы и прекращение транскрипции. В этот момент полимераза располагается на полиурациловом участке последовательности. Слабые аденин-урациловые связи снижают энергию дестабилизации дуплекса РНК-ДНК, что позволяет им ослабить напряжение полинуклеотидной цепи и диссоциировать от РНК-полимеразы.
Белок контрольной точки клеточного цикла RAD17 — белок, кодируемый у человека геном RAD17 .
ATM — серин/треониновая протеинкиназа, которая рекрутируется и активируется двунитевыми разрывами ДНК. Эта киназа фосфорилирует несколько ключевых белков, которые инициируют остановку клеточного цикла, запускают репарацию ДНК или апоптоз. Некоторые из этих белков, в том числе p53, Сhk2 и вариантный гистон H2AX являются опухолевыми супрессорами. Белок кодируется геном ATM.

ДНК (цитозин-5)-метилтрансфераза 3-подобный фермент англ. DNA (cytosine-5)-methyltransferase 3-like, DNMT3L — фермент, кодируемый у человека геном DNMT3L.

Cas9 — это управляемая при помощи РНК-гидов эндонуклеаза, связанная с адаптивной иммунной системой CRISPR у ряда бактерий, в частности Streptococcus pyogenes. S. pyogenes использует Cas9 для запоминания, последующей проверки и разрезания чужеродной ДНК, например, ДНК бактериофагов или плазмид.
Геномная нестабильность определяется высокой частотой мутаций в геноме клеточной линии. Эти мутации могут включать в себя изменения в последовательности нуклеиновых кислот, хромосомные перестройки или анеуплоидию. Геномная нестабильность является центральным фактором канцерогенеза, но также фактором некоторых нейродегенеративных заболеваний, таких как боковой амиотрофический склероз или нервно-мышечное заболевание миотоническая дистрофия.
Белок эксцизионной репарации ДНК ERCC-6 — белок, кодируемый у человека геном ERCC6 . Ген ERCC6 расположен на длинном плече хромосомы 10 в позиции 11.23.

Нибрин, также известный как NBN или Nbs1 — белок, кодируемый у человека геном NBN.
Белок репарации ДНК RAD50, также известный как RAD50 — белок, кодируемый у человека геном RAD50.

Белок репарации двойных разрывов нитей MRE11A — белок, кодируемый у человека геном MRE11A.

Негомологи́чное соедине́ние концо́в, или негомологи́чное воссоедине́ние концо́в — один из путей репарации двунитевых разрывов в ДНК. Негомологичным этот процесс называется потому, что повреждённые концы цепи соединяются лигазой напрямую, не нуждаясь в гомологичном шаблоне, в отличие от процесса гомологичной рекомбинации. NHEJ существенно менее точен, чем гомологичная рекомбинация, и часто он приводит к потере нуклеотидов, транслокациям или слиянию теломер, при этом последние два могут являться признаками опухолевой клетки. NHEJ обнаружен у представителей всех царств живой природы, кроме того, в клетках млекопитающих он служит основным путём репарации двуцепочечных разрывов.
Микрогомологичное соединение концов (МСК), также известное как альтернативное негомологичное соединение концов (Альт-НСК), является одним из путей репарации двунитевых разрывов в ДНК-цепи. Как рассмотрено Маквеем и Ли, главным отличительным свойством МСК является использование микрогомологичных последовательностей, состоящих из 5-25 пар оснований (п.о.) МСК часто связано с хромосомными аномалиями, такими как делеции, транслокации, инверсии и другие сложные перестройки.

Трёхцепочечная ДНК, H-ДНК или три́плекс-ДНК — форма ДНК, в которой три олигонуклеотида обвивают друг друга, формируя тройную спираль. В трёхцепочечной ДНК третья цепь ДНК связывается с двуцепочечной B-формой ДНК, сформированной за счёт уотсон—криковских взаимодействий, с помощью хугстиновских взаимодействий или обратных хугстиновских водородных связей. Трёхцепочечная ДНК может мешать нормальной репликации и повышать частоту мутаций в области формирования.
CRISPR-Cas13 это РНК-нуклеаза которую можно использовать для целенаправленной деградации РНК. Ферменты Cas13 имеют два эндоРНКазных домена HEPN. Для нацеливания этот фермент использует направляющую РНК (gRNA). После активации путем спаривания между последовательностью CRISPR РНК (crРНК) и комплементарной одноцепочечной РНК (ssРНК)-мишенью эффектор Cas13 вызывает расщепление РНК-мишени, При этом для активации рибонуклеазы Cas13 требуется почти идеальная комплементарность между целевой РНК и Cas13-ассоциированной направляющей РНК. Поскольку программируемая последовательность направляющей РНК Cas13 примерно в три раза больше, чем исходная последовательность кшРНК, опосредованный CRISPR-Cas13 сайленсинг транскрипции дает исследователям большую избирательность цели. Еще одним преимуществом является то, что Cas13 не требует последовательности PAM, что теоретически позволяет ему нацеливаться практически на любую область РНК.
![Димерная рибонуклеаза T (зелёным и синим цветами показаны отдельные мономеры фермента) в комплексе с ДНК (показана оранжевым цветом), PDB ID 3NH1.[1]](https://upload.wikimedia.org/wikipedia/commons/thumb/0/01/RnaseT.png/274px-RnaseT.png)