Сайт связывания

В биохимии и молекулярной биологии сайт связывания (участок связывания) — это участок макромолекулы, такой как белок, который специфично связывается с другой молекулой[1]. Партнер связывания макромолекулы часто называют лигандом[2]. Лиганды могут включать другие белки (приводящие к взаимодействию белок-белок)[3][4], ферментные субстраты[5], вторичные мессенджеры, гормоны или аллостерические модуляторы[6]. Событие связывания часто, но не всегда, сопровождается конформационным изменением, которое изменяет функцию белка[7]. Связывание с сайтами связывания белков чаще всего является обратимым (временным и нековалентным), но также может быть ковалентным обратимым[8] или необратимым[9].
Функция
Связывание лиганда с сайтом связывания на белке часто вызывает изменение конформации белка и приводит к изменению клеточной функции. Следовательно, сайты связывания на белке являются критическими частями путей передачи сигнала[10]. Типы лигандов включают нейротрансмиттеры, токсины, нейропептиды и стероидные гормоны[11]. Сайты связывания подвергаются функциональным изменениям в ряде контекстов, включая ферментативный катализ, передачу сигналов молекулярного пути, гомеостатическую регуляцию и физиологическую функцию. Электрический заряд, стерическая форма и геометрия сайта избирательно позволяют связывать высокоспецифичные лиганды, активируя определённый каскад клеточных взаимодействий, за которые отвечает белок[12][13].
Катализ

Ферменты вызывают катализ, более прочно связываются с переходными состояниями, чем субстраты и продукты. В сайте каталитического связывания на субстрат могут действовать несколько различных взаимодействий. Они варьируются от электрокатализа, кислотного и основного катализа до ковалентного катализа и катализа ионами металлов[11]. Эти взаимодействия уменьшают энергию активации химической реакции, обеспечивая благоприятные взаимодействия для стабилизации высокоэнергетической молекулы. Связывание ферментов обеспечивает более близкое расположение и исключение веществ, не имеющих отношения к реакции. Это специфическое связывание также препятствует побочным реакциям[14][11].
Типы ферментов, которые могут выполнять эти действия, включают оксидоредуктазы, трансферазы, гидролазы, лиазы, изомеразы и лигазы[15].
Например, гексокиназа трансферазы катализирует фосфорилирование глюкозы с образованием глюкозо-6-фосфата. Остатки активного центра гексокиназы позволяют стабилизировать молекулу глюкозы в активном центре и стимулируют начало альтернативного пути благоприятных взаимодействий, снижая энергию активации[16].
Ингибирование
Ингибирование белка связыванием ингибитора может вызвать нарушение регуляции пути, гомеостатической регуляции и физиологической функции.
Конкурентные ингибиторы конкурируют с субстратом за связывание со свободными ферментами в активных центрах и, таким образом, препятствуют образованию комплекса фермент-субстрат при связывании. Например, отравление оксидом углерода вызвано конкурентным связыванием оксида углерода, в отличие от кислорода в гемоглобине.
В качестве альтернативы неконкурентные ингибиторы связываются одновременно с субстратом в активных центрах. При связывании с комплексом ферментного субстрата (ES) образуется комплекс ингибитора ферментного субстрата (ESI). Подобно конкурентным ингибиторам, также снижается скорость образования продукта[5].
Наконец, смешанные ингибиторы способны связываться как со свободным ферментом, так и с комплексом фермент-субстрат. Однако, в отличие от конкурентных и неконкурентных ингибиторов, смешанные ингибиторы связываются с аллостерическим сайтом. Аллостерическое связывание вызывает конформационные изменения, которые могут увеличивать сродство белка к субстрату. Это явление называется положительной модуляцией. И наоборот, аллостерическое связывание, которое снижает сродство белка к субстрату, является отрицательной модуляцией[17].
Типы
Активный центр
В активном центре субстрат связывается с ферментом, вызывая химическую реакцию[18][19]. Субстраты, переходные состояния и продукты могут связываться с активным сайтом, как и любые конкурентные ингибиторы[18]. Например, в контексте функции белков связывание кальция с тропонином в мышечных клетках может вызывать конформационные изменения тропонина. Это позволяет тропомиозину открывать сайт связывания актин-миозин, с которым связывается миозиновая головка, чтобы образовать поперечный мостик и вызвать сокращение мышц[20].
В контексте крови примером конкурентного связывания является окись углерода, которая конкурирует с кислородом за активный центр гема. Высокое сродство окиси углерода может превосходить кислород в присутствии низкой концентрации кислорода. В этих обстоятельствах связывание монооксида углерода вызывает изменение конформации, которое препятствует связыванию гема с кислородом, что приводит к отравлению монооксидом углерода[5].

Аллостерический сайт
В регуляторном сайте связывание лиганда может вызывать усиление или ингибирование функции белка[5][21]. Связывание лиганда с аллостерическим сайтом мультимерного фермента часто вызывает положительную кооперативность, то есть связывание одного субстрата вызывает благоприятное изменение конформации и увеличивает вероятность связывания фермента со вторым субстратом[22]. Лиганды регуляторного сайта могут включать гомотропные и гетеротропные лиганды, в которых один или несколько типов молекул влияют на активность фермента соответственно[23].
Регулируемые ферменты часто играют важную роль в метаболических путях. Например, фосфофруктокиназа (ФФК), которая фосфорилирует фруктозу при гликолизе, в значительной степени регулируется АТФ. Его регуляция в гликолизе является обязательной, потому что это этап метаболизма, ограничивающий скорость. ФФК также контролирует количество глюкозы, предназначенной для образования АТФ через катаболический путь. Следовательно, при достаточном уровне АТФ, ФФК аллостерически ингибируется АТФ. Это регулирование эффективно сохраняет запасы глюкозы, которые могут потребоваться для других путей. Цитрат, промежуточный продукт цикла лимонной кислоты, также работает как аллостерический регулятор ФФК[23][24].
Одно- и многоцепочечные сайты связывания
Сайты связывания можно охарактеризовать также по их структурным особенностям. Одноцепочечные сайты («монодесмических» лигандов, μόνος: одиночные, δεσμός: связывание) образованы одной белковой цепью, в то время как многоцепочечные сайты («полидесмических» лигандов, πολοί: многие)[25] часто встречаются в белковых комплексах и образуются лигандами, которые связывают более одной белковой цепи, как правило, на границах раздела белков или рядом с ними. Недавние исследования показывают, что структура сайта связывания имеет сильные последствия для биологии белковых комплексов (эволюция функции, аллостерия)[26][27].
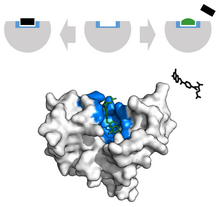
Скрытые сайты связывания
Скрытые сайты связывания представляют собой сайты связывания, которые временно образуются в форме «апо» или индуцируются связыванием лиганда. Принятие во внимание скрытых сайтов связывания увеличивает размер протеома человека, потенциально «поддающегося лекарству», с ~40 % до ~78 % белков, связанных с заболеванием[28]. Сайты связывания были исследованы с помощью: машины опорных векторов, примененной к набору данных «CryptoSite»[28], расширения набора данных «CryptoSite»[29], моделирования молекулярной динамики в долгосрочном масштабе с использованием модели состояния Маркова и с помощью биофизических экспериментов[30], и индекса скрытых сайтов, основанного на относительной доступной площади поверхности[31].
Кривые привязки

Кривые связывания описывают процесс связывания лиганда с белком. Кривые могут быть охарактеризованы их формой, сигмоидальной или гиперболической, которая отражает, проявляет ли белок кооперативное или некооперативное поведение связывания соответственно[32]. Обычно ось абсцисс описывает концентрацию лиганда, а ось ординат описывает фракционное насыщение лигандов, связанных со всеми доступными сайтами связывания[5]. Уравнение Михаэлиса Ментен обычно используется при определении формы кривой. Уравнение Михаэлиса Ментен выводится на основе стационарных условий и учитывает ферментативные реакции, протекающие в растворе. Однако, когда реакция происходит, когда фермент связан с субстратом, кинетика развивается по-другому[33].
Моделирование с помощью кривых связывания полезно при оценке аффинности связывания кислорода с гемоглобином и миоглобином в крови. Гемоглобин, который имеет четыре группы гема, проявляет кооперативное связывание. Это означает, что связывание кислорода с гемовой группой на гемоглобине вызывает благоприятное изменение конформации, которое позволяет повысить благоприятность связывания кислорода для следующих гемовых групп. В этих обстоятельствах кривая связывания гемоглобина будет сигмоидальной из-за его повышенной способности связываться с кислородом. Поскольку миоглобин имеет только одну гемовую группу, он демонстрирует некооперативное связывание, которое является гиперболическим на кривой связывания[34].
Практическое применение
Биохимические различия между разными организмами и людьми полезны для разработки лекарств. Например, пенициллин ингибирует бактериальные ферменты DD-транспептидазы, нарушая синтез бактериальной клеточной стенки и вызывая гибель клеток. Таким образом, изучение сайтов связывания актуально для многих областей исследований, включая механизмы рака[7], лекарственные формы[35] и физиологическую регуляцию[36]. Разработка ингибиторов для подавления функции белка является распространенной формой фармацевтической терапии[37].
В области лечения рака лиганды, которые отредактированы, чтобы иметь внешний вид, подобный природному лиганду, используются для подавления роста опухоли. Например, химиотерапевтический метотрексат действует как конкурентный ингибитор активного центра дигидрофолатредуктазы[38]. Это взаимодействие подавляет синтез тетрагидрофолата, останавливая производство ДНК, РНК и белков[38]. Ингибирование этой функции подавляет рост опухолей и улучшает состояние при тяжелом псориазе и ревматоидном артрите у взрослых[37].
При сердечно-сосудистых заболеваниях для лечения пациентов с гипертонией используются такие препараты, как бета-блокаторы. Бета-блокаторы (β-блокаторы) — это антигипертензивные средства, которые блокируют связывание гормонов адреналина и норадреналина с рецепторами β1 и β2 в сердце и кровеносных сосудах. Эти рецепторы обычно опосредуют симпатическую реакцию «бей или беги», вызывая сужение кровеносных сосудов[39].
Конкурентные ингибиторы также широко распространены в коммерции. Ботулинический токсин, известный под коммерческим названием Ботокс, представляет собой нейротоксин, вызывающий вялый паралич в мышцах из-за связывания с ацетилхолинзависимыми нервами. Это взаимодействие подавляет мышечные сокращения, создавая вид гладкой мускулатуры[40].
Прогнозирование
Для прогнозирования расположения сайтов связывания на белках был разработан ряд вычислительных инструментов[21][41][42]. Они могут быть в широком смысле классифицированы на основе последовательности или структуры[42]. Методы, основанные на последовательности, основаны на предположении, что последовательности функционально сохраненных частей белков, таких как сайт связывания, консервативны. Методы, основанные на структуре, требуют трехмерной структуры белка. Эти методы, в свою очередь, могут быть подразделены на шаблонные и «карманные» методы[42]. Методы, основанные на шаблонах, ищут 3D-сходство между целевым белком и белками с известными сайтами связывания. Методы, основанные на карманах, ищут вогнутые поверхности или скрытые карманы в целевом белке, которые обладают такими характеристиками, как гидрофобность и способность связывать водород, что позволило бы им связывать лиганды с высоким сродством[42]. Несмотря на то, что здесь используется термин «карман», аналогичные методы могут быть использованы для прогнозирования сайтов связывания, используемых в белково-белковых взаимодействиях, которые обычно являются более плоскими, а не в «карманах»[43].
Примечания
- ↑ Binding site. Medical Subject Headings (MeSH). U.S. National Library of Medicine. — «The parts of a macromolecule that directly participate in its specific combination with another molecule.» Дата обращения: 14 августа 2021. Архивировано 17 июня 2020 года.
- ↑ Ligands. Medical Subject Headings (MeSH). U.S. National Library of Medicine. — «A molecule that binds to another molecule, used especially to refer to a small molecule that binds specifically to a larger molecule.» Дата обращения: 14 августа 2021. Архивировано 17 июня 2020 года.
- ↑ "Binding site prediction for protein-protein interactions and novel motif discovery using re-occurring polypeptide sequences". BMC Bioinformatics. 12: 225. June 2011. doi:10.1186/1471-2105-12-225. PMID 21635751.
{{cite journal}}: Википедия:Обслуживание CS1 (не помеченный открытым DOI) (ссылка) - ↑ "3D U-Net: A Voxel-based method in binding site prediction of protein structure". Journal of Bioinformatics and Computational Biology. 19 (1): 1—10. April 2021. doi:10.1142/S0219720021500062. ISSN 1757-6334.
- ↑ 1 2 3 4 5 Charles Hardin. Biochemistry : essential concepts. — New York, 2013. — 1 online resource (xviii, 316 pages) с. — ISBN 978-1-62870-176-0, 1-62870-176-5.
- ↑ Allosteric receptor modulation in drug targeting. — New York: Taylor & Francis, 2006. — 1 online resource (xii, 359 pages, 1 unnumbered leaf of plates) с. — ISBN 978-1-4200-1618-5, 1-4200-1618-0, 0-8247-2791-6, 978-0-8247-2791-8, 1-280-86341-2, 978-1-280-86341-7, 9786610863419, 6610863415.
- ↑ 1 2 "Protein function annotation by local binding site surface similarity". Proteins. 82 (4): 679—94. April 2014. doi:10.1002/prot.24450. PMID 24166661.
- ↑ "Targeting biomolecules with reversible covalent chemistry". Current Opinion in Chemical Biology. 34: 110—116. October 2016. doi:10.1016/j.cbpa.2016.08.011. PMID 27599186.
- ↑ Andrea Bellelli. Reversible ligand binding : theory and experiment. — First edition. — Hoboken, NJ, 2018. — 1 online resource (xiii, 289 pages) с. — ISBN 978-1-119-23850-8, 1-119-23850-1, 978-1-119-23847-8, 1-119-23847-1, 978-1-119-23849-2, 1-119-23849-8, 1-119-23848-X, 978-1-119-23848-5.
- ↑ "Small-molecule binding sites to explore protein-protein interactions in the cancer proteome". Molecular BioSystems. 12 (10): 3067—87. October 2016. doi:10.1039/c6mb00231e. PMID 27452673.
- ↑ 1 2 3 Principles and techniques of biochemistry and molecular biology.. — [Place of publication not identified], 2009. — 1 online resource с. — ISBN 0-511-84147-7, 978-0-511-84147-7.
- ↑ Biochemistry Free For All. — Oregon State University, 2015. — P. 110–141.
- ↑ "Allosteric binding sites in Rab11 for potential drug candidates". PLOS One. 13 (6): e0198632. 2018-06-06. doi:10.1371/journal.pone.0198632. PMID 29874286.
{{cite journal}}: Википедия:Обслуживание CS1 (не помеченный открытым DOI) (ссылка) - ↑ C. M. Dobson. Foundations of chemical biology. — Oxford [England], 2001. — [ii], 97 pages с. — ISBN 0-19-924899-0, 978-0-19-924899-5.
- ↑ Polymer and Biopolymer Brushes. — 2017-12-04. — ISBN 978-1-119-45501-1. — doi:10.1002/9781119455042.
- ↑ Dictionary of Food Science and Technology (2nd Edition). — International Food Information Service, 2009. — ISBN 978-1-4051-8740-4.
- ↑ Bioprocess engineering. — Woodhead Publishing. — P. 79–84. — ISBN 978-1-78242-167-2.
- ↑ 1 2 Enzymes // Principles and Techniques of Biochemistry and Molecular Biology : [англ.]. — Cambridge University Press, March 2010. — P. 581–624. — ISBN 9780511841477. — doi:10.1017/cbo9780511841477.016.
- ↑ Dictionary of Chemical Engineering. — Oxford University Press. — ISBN 978-1-62870-844-8.
- ↑ Biology How Life Works. — W.H. Freeman and Company. — P. 787–792. — ISBN 978-1-4641-2609-3.
- ↑ 1 2 "Binding site comparison for function prediction and pharmaceutical discovery". Current Opinion in Structural Biology. 25: 34—9. April 2014. doi:10.1016/j.sbi.2013.11.012. PMID 24878342.
- ↑ Prokaryotic Intercellular Signalling. — 2004. — ISBN 9789048164837. — doi:10.1007/978-94-017-0998-9_2.
- ↑ 1 2 The Biophysical Chemistry of Nucleic Acids & Proteins. — 2010. — ISBN 978-0956478115.
- ↑ Biotechnological Innovations in Chemical Synthesis. — Butterworth-Heinemann, 1997. — ISBN 978-0-7506-0561-8.
- ↑ "Ligand Binding Site Structure Shapes Folding, Assembly and Degradation of Homomeric Protein Complexes". Journal of Molecular Biology. 431 (19): 3871—3888. 2019. doi:10.1016/j.jmb.2019.07.014. PMID 31306664.
- ↑ "Ligand Binding Site Structure Influences the Evolution of Protein Complex Function and Topology". Cell Reports. 22 (12): 3265—3276. 2018. doi:10.1016/j.celrep.2018.02.085. PMID 29562182.
- ↑ "Ligand-Binding-Site Structure Shapes Allosteric Signal Transduction and the Evolution of Allostery in Protein Complexes". Molecular Biology and Evolution. 36 (8): 1711—1727. 2019. doi:10.1093/molbev/msz093. PMID 31004156.
- ↑ 1 2 "CryptoSite: Expanding the Druggable Proteome by Characterization and Prediction of Cryptic Binding Sites". Journal of Molecular Biology. 428 (4): 709—719. February 2016. doi:10.1016/j.jmb.2016.01.029. PMID 26854760.
- ↑ "Exploring the structural origins of cryptic sites on proteins". Proceedings of the National Academy of Sciences of the United States of America. 115 (15): E3416–E3425. April 2018. doi:10.1073/pnas.1711490115. PMID 29581267.
- ↑ "Discovery of multiple hidden allosteric sites by combining Markov state models and experiments". Proceedings of the National Academy of Sciences of the United States of America. 112 (9): 2734—9. March 2015. doi:10.1073/pnas.1417811112. PMID 25730859.
- ↑ "Structural Fluctuations of Aromatic Residues in an Apo-Form Reveal Cryptic Binding Sites: Implications for Fragment-Based Drug Design". The Journal of Physical Chemistry B. 124 (45): 9977—9986. November 2020. doi:10.1021/acs.jpcb.0c04963.
- ↑ "Teaching biochemistry online at Oregon State University". Biochemistry and Molecular Biology Education. 45 (1): 25—30. January 2017. doi:10.1002/bmb.20979. PMID 27228905.
- ↑ "Kinetics of enzyme action on surface-attached substrates: a practical guide to progress curve analysis in any kinetic situation". Langmuir. 28 (41): 14665—71. October 2012. doi:10.1021/la3030827. PMID 22978617.
- ↑ Biology: how life works. — ISBN 9781464126093.
- ↑ "Apolipoprotein A-IV: A potential therapeutic target for atherosclerosis". Prostaglandins & Other Lipid Mediators. 139: 87—92. November 2018. doi:10.1016/j.prostaglandins.2018.10.004. PMID 30352313.
- ↑ "Skeletal myosin binding protein-C: An increasingly important regulator of striated muscle physiology". Archives of Biochemistry and Biophysics. 660: 121—128. December 2018. doi:10.1016/j.abb.2018.10.007. PMID 30339776.
- ↑ 1 2 "Understanding and managing methotrexate nephrotoxicity". The Oncologist. 11 (6): 694—703. June 2006. doi:10.1634/theoncologist.11-6-694. PMID 16794248.
- ↑ 1 2 "Interaction of dihydrofolate reductase with methotrexate: ensemble and single-molecule kinetics". Proceedings of the National Academy of Sciences of the United States of America. 99 (21): 13481—6. October 2002. doi:10.1073/pnas.172501499. PMID 12359872.
- ↑ Current Cardiovascular Drugs. — 2000. — ISBN 978-1-57340-135-7.
- ↑ "Botulinal neurotoxins: revival of an old killer". Current Opinion in Pharmacology. 5 (3): 274—9. June 2005. doi:10.1016/j.coph.2004.12.006. PMID 15907915.
- ↑ "Proteins and Their Interacting Partners: An Introduction to Protein-Ligand Binding Site Prediction Methods". International Journal of Molecular Sciences. 16 (12): 29829—42. December 2015. doi:10.3390/ijms161226202. PMID 26694353.
{{cite journal}}: Википедия:Обслуживание CS1 (не помеченный открытым DOI) (ссылка) - ↑ 1 2 3 4 "Can We Rely on Computational Predictions To Correctly Identify Ligand Binding Sites on Novel Protein Drug Targets? Assessment of Binding Site Prediction Methods and a Protocol for Validation of Predicted Binding Sites". Cell Biochemistry and Biophysics. 75 (1): 15—23. March 2017. doi:10.1007/s12013-016-0769-y. PMID 27796788.
- ↑ "Analysis of protein-protein interaction sites using surface patches". Journal of Molecular Biology. 272 (1): 121—32. September 1997. doi:10.1006/jmbi.1997.1234. PMID 9299342.
Ссылки
| Активность | |
|---|---|
| Регуляция | |
| Классификация | |
| Типы |
|