
Кроссинго́вер — процесс обмена участками гомологичных хромосом во время конъюгации в профазе первого деления мейоза, которое происходит, например, при образовании гамет или спор. Помимо мейотического, описан также митотический кроссинговер.
Дрейф ге́нов, или гене́тико-автомати́ческие проце́ссы, или эффект Райта — случайные изменения частот аллелей и генотипов, происходящие в небольшой полиморфной популяции при смене поколений. Такие случайные изменения приводят к двум характерным последствиям: сначала к флуктуациям частоты аллеля из поколения в поколение, а в конце концов к полному закреплению или элиминации данного аллеля. Влияние дрейфа генов на изменение частот аллелей в данной популяции зависит прежде всего от её размеров.
Сцепленное наследование — скоррелированное наследование определённых состояний генов, расположенных в одной хромосоме.
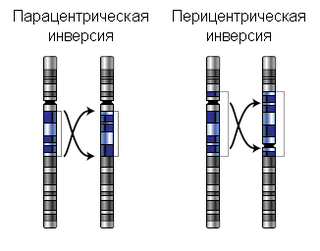
Инве́рсия — хромосомная перестройка, при которой происходит поворот участка хромосомы на 180°. Инверсии являются сбалансированными внутрихромосомными перестройками. Различают парацентрические и перицентрические инверсии. Инверсии играют роль в эволюционном процессе, видообразовании и в нарушениях фертильности.

Y-хромосо́ма — одна из двух половых хромосом в системе хромосомного определения пола XY, которая встречается у многих животных, в том числе у большинства млекопитающих, включая человека. У млекопитающих содержит ген SRY, определяющий мужской пол организма, а также гены, необходимые для нормального формирования сперматозоидов. Мутации в гене SRY могут привести к формированию женского организма с генотипом XY. Y-хромосома человека состоит из более чем 62 миллионов пар нуклеотидов.
Эволюционная теория номадических генов была предложена В. Геодакяном в 1996 г. Так же как и эволюционные теории пола и асимметрии, номадическая теория генов основана на принципе сопряженных подсистем, которые эволюционируют асинхронно. Теории пола и половых хромосом тесно взаимосвязаны и описывают процесс эволюции признака на разных уровнях организации. В филогенезе эволюция у мужского пола начинается и кончается раньше, чем у женского поскольку признаки появляются сначала в генотипе мужских особей, и только потом, через много поколений, передаются женским. Половые хромосомы непосредственно участвуют в этом процессе. Ещё в 1965 году В. Геодакян предположил, что «…в хромосомном наборе половые хромосомы выполняют роль оперативной памяти, а аутосомы — постоянной, поэтому половые хромосомы являются „воротами“ для изменчивости в наследственность». Позднее, в ряде работ было показано, что частота мутаций в Y-хромосоме действительно выше, чем в других хромосомах.
Парасексуа́льный проце́сс, или парасексуа́льный цикл, — аналог полового процесса, свойственный грибам и одноклеточным организмам. При парасексуальном процессе происходит слияние вегетативных клеток, содержащих генетически разнородные ядра, возникновение гетерозиготных диплоидов за счёт слияния ядер в гетерокарионах и последующего митотического расщепления диплоидов, приводящее к появлению гаплоидных или диплоидных рекомбинантов.

Си́напсис — конъюгация хромосом, попарное временное сближение гомологичных хромосом, во время которого между ними может произойти обмен гомологичными участками. Происходит в профазе I мейоза. Когда гомологичные хромосомы синаптируют, их концы прикрепляются к ядерной оболочке. Потом такие концевые мембранные комплексы перемещаются при содействии ядерного цитоскелета, пока соответственные концы хромосом не объединятся в пары. После этого и межконцевые участки хромосом начинают сближаться, при этом они могут соединяться посредством РНК-белкового комплекса, называемого синаптонемальным комплексом. Синапсису подвергаются только аутосомы в процессе мейоза, половые же хромосомы остаются неспаренными.
Хромосомные перестройки — тип мутаций, которые изменяют структуру хромосом. Классифицируют следующие виды хромосомных перестроек: делеции, инверсии, дупликации, транслокации, а также дицентрические и кольцевые хромосомы. Известны также изохромосомы, несущие два одинаковых плеча. Если перестройка изменяет структуру одной хромосомы, то такую перестройку называют внутрихромосомной, если же двух разных, то межхромосомной. Хромосомные перестройки подразделяют также на сбалансированные и несбалансированные. Сбалансированные перестройки не приводят к потере или добавлению генетического материала при формировании, поэтому их носители, как правило, фенотипически нормальны. Несбалансированные перестройки меняют дозовое соотношение генов, и, как правило, их носительство сопряжено с существенными отклонениями от нормы.
Синдро́м де ля Шапе́ля — назван в честь исследователя, впервые охарактеризовавшего его в 1972 году. Синдром относится к редкой хромосомной патологии, возникающей в результате кроссинговера между X- и Y-хромосомами в процессе мейоза, в результате чего одна или обе X-хромосомы содержат нормальный мужской ген SRY. Распространённость данного синдрома составляет 4—5 на 100 000, что меньше встречаемости синдрома Клайнфельтера.

Генети́ческая ка́рта — схема взаимного расположения структурных генов, регуляторных элементов и генетических маркеров, а также относительных расстояний между ними на хромосоме. Метод построения генетических карт называется генетическим картированием.

Хиа́зма — в генетике точка, в которой две гомологичные несестринские хроматиды обмениваются генетическим материалом в ходе кроссинговера в течение мейоза. Хиазмы становятся видны в фазе диплотены профазы I мейоза, но сам кроссинговер происходит в предыдущей фазе — пахитене. Когда каждая тетрада, состоящая из двух пар сестринских хроматид, начинает распадаться, хиазмы остаются единственными точками контакта.
Митоти́ческий кроссинго́вер — тип генетической рекомбинации, который может проходить в соматических клетках при митотических делениях как у организмов, обладающих полом, так и бесполых организмов. В случае бесполых организмов митотическая рекомбинация является единственным ключом к пониманию сцепления генов, так как у таких организмов это единственный способ генетической рекомбинации. Кроме того, митотическая рекомбинация может привести к мозаичной экспрессии рецессивных признаков у гетерозиготной особи. Такая экспрессия имеет важное значение в онкогенезе, она также позволяет изучать летальные рецессивные мутации.
Тетра́да — четыре споры, образовавшиеся после ряда делений и рекомбинации у некоторых грибов и одноклеточных водорослей. Если два родителя различаются по двум аллелям двух разных генов, то возможные споры в тетраде можно подразделить на три типа: родительский дитип (Р), неродительский дитип (N) и тетратип (T).
Синаптоне́мный ко́мплекс (СК) — белковый комплекс, формирующийся между гомологичными хромосомами в ходе мейоза. Синаптонемный комплекс предположительно является связующим звеном между хромосомами во время спаривания (синапсиса).

Хромосомная теория наследственности — теория, согласно которой передача наследственной информации в ряду поколений связана с передачей хромосом, в которых в определённой и линейной последовательности расположены гены. Эта теория сформулирована в начале XX века, основной вклад в её создание внесли американский цитолог У. Саттон, немецкий эмбриолог Т. Бовери и американский генетик Т. Морган со своими сотрудниками К. Бриджесом, А. Стёртевантом и Г. Мёллером.

Микрохромосо́мы — очень маленькие хромосомы, типичные для кариотипов птиц, некоторых рептилий, рыб и земноводных; у млекопитающих они, судя по всему, отсутствуют. Их размер составляет меньше 20 мегабаз; хромосомы, чей размер превышает 40 мегабаз, называются макрохромосомами, а хромосомы размером от 20 до 40 мегабаз — промежуточными хромосомами.

Структу́ра Холлиде́я — структура из четырёх цепей нуклеиновых кислот, соединённых друг с другом водородными связями с образованием четырёх двуцепочечных ветвей. Эти ветви могут принимать несколько различных конформаций в зависимости от концентрации солей в окружающем буферном растворе и последовательности нуклеотидов, располагающихся в непосредственной близости от точки соединения. Структура названа в честь английского молекулярного биолога Робина Холлидея, который предположил её существование в 1964 году.

Гомологи́чная рекомбина́ция, или о́бщая рекомбина́ция, — тип генетической рекомбинации, во время которой происходит обмен нуклеотидными последовательностями между двумя похожими или идентичными хромосомами. Это наиболее широко используемый клетками способ устранения двух- или однонитевых повреждений ДНК. Гомологичная рекомбинация также создаёт разнообразие комбинаций генов во время мейоза, обеспечивающих высокий уровень наследственной изменчивости, что, в свою очередь, позволяет популяции лучше адаптироваться в ходе эволюции. Различные штаммы и виды бактерий и вирусов используют гомологичную рекомбинацию в процессе горизонтального переноса генов.
Балансёрная хромосома — элемент генетического инструментария, специально модифицированная хромосома, используемая для поддержания некоторого гена в искусственной популяции в гетерозиготном состоянии и быстрого поиска подобных особей. Также используется для компенсации повреждённого гена и предотвращения рекомбинации (кроссинговера) между гомологичными хромосомами в мейозе. Наиболее часто используется в генетике Drosophila melanogaster, что позволяет поддерживать в популяции мутацию в состоянии гетерозиготы без постоянного контроля её наличия; также может использоваться для мышей. Балансерная хромосома имеет три важных свойства: она подавляет рекомбинацию с гомологичной хромосомой, несёт в себе доминантные маркеры, негативно влияет на репродуктивную функцию в состоянии гомозиготы.
![{\displaystyle P\left[{\text{рекомбинация}}|{\text{сцепленность }}d{\text{ cM}}\right]=\sum _{k=0}^{\infty }P\left[2k+1{\text{ кроссинговеров}}|{\text{сцепленность }}d{\text{ cM}}\right]=}](https://wikimedia.org/api/rest_v1/media/math/render/svg/1efaed1b49439e712602c3561cfcd8549ed3d907)

![{\displaystyle d=50\ln \left({\frac {1}{1-2P\left[{\text{рекомбинация}}\right]}}\right)\,.}](https://wikimedia.org/api/rest_v1/media/math/render/svg/8abd4a86bc4736ee99545bf3dfdbe05677eecdb0)