
Гипо́физ — мозговой придаток в форме округлого образования, расположенного на нижней поверхности головного мозга в костном кармане, называемом турецким седлом, вырабатывает гормоны, влияющие на рост, обмен веществ и репродуктивную функцию. Является центральным органом эндокринной системы; тесно связан и взаимодействует с гипоталамусом.

Промежуточный мозг, или диэнцефалон — отдел головного мозга хордовых животных, который образуется в процессе эмбрионального развития из задней части зародышевого переднего мозга (прозэнцефалона). На пятипузырьковой стадии из задней части зародышевого переднего мозга (прозэнцефалона) образуется вторичный мозговой пузырь — зародышевый промежуточный мозг, или зародышевый диэнцефалон. В то же время из передней части зародышевого переднего мозга образуется другой вторичный мозговой пузырь — зародышевый конечный мозг, или зародышевый телэнцефалон. Во взрослом состоянии структуры промежуточного мозга располагаются по бокам (латеральнее) третьего желудочка головного мозга.

Кортикотропин-рилизинг-гормон, или кортикорелин, кортиколиберин, кортикотропин-рилизинг-фактор, сокращённо КРГ, — один из представителей класса рилизинг-гормонов гипоталамуса. Он действует на переднюю долю гипофиза и вызывает там секрецию АКТГ.
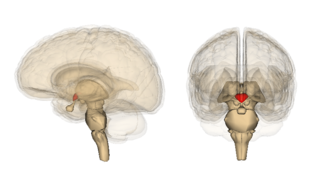
Гипотала́мус — небольшая область в промежуточном мозге, включающая в себя большое число групп клеток, которые регулируют нейроэндокринную деятельность мозга и гомеостаз организма. Гипоталамус связан нервными путями практически со всеми отделами центральной нервной системы, включая кору, гиппокамп, миндалину, мозжечок, ствол мозга и спинной мозг. Вместе с гипофизом гипоталамус образует гипоталамо-гипофизарную систему, в которой гипоталамус управляет выделением гормонов гипофиза и является центральным связующим звеном между нервной и эндокринной системами. Он выделяет гормоны и нейропептиды и регулирует такие функции, как ощущение голода и жажды, терморегуляция организма, половое поведение, сон и бодрствование. Исследования последних лет показывают, что гипоталамус играет важную роль и в регуляции высших функций, таких как память и эмоциональное состояние, и тем самым участвует в формировании различных аспектов поведения.
Соматотропин-рилизинг-гормон, или соматрелин, соматолиберин, соматотропин-рилизинг-фактор, сокращённо СРГ или СРФ — один из представителей класса рилизинг-гормонов гипоталамуса.

База́льные я́дра — несколько скоплений серого вещества, расположенных в белом веществе латеральнее таламуса на уровне основания полушарий конечного мозга. Базальные ядра входят в состав переднего мозга, расположенного на границе между лобными долями и над стволом мозга. Традиционно в состав базальных ядер включались полосатое тело, в свою очередь состоящее из хвостатого ядра, скорлупы и бледного шара, а также ограда и миндалевидное тело. Бледный шар и скорлупа вместе называются чечевицеобразным ядром. Белое вещество между таламусом и чечевицеобразным ядром называется внутренней капсулой, между чечевицеобразным ядром и оградой — наружной капсулой и между оградой и островком — крайней капсулой. Эта классификация основана на топографии анатомического среза мозга, однако в последнее время она всё чаще заменяется функциональной, где под термином «базальные ядра» понимают полосатое тело и несколько ядер промежуточного и среднего мозга, которые совместно обеспечивают функциональную регуляцию движений и мотивационных аспектов поведения. Функции ограды остаются недостаточно изученными, а структуры миндалевидного тела относят к лимбической системе.

Вентральная область покрышки — часть среднего мозга, расположенная дорсомедиально по отношению к чёрному веществу и вентрально по отношению к красным ядрам. Является началом мезокортикального и мезолимбического дофаминовых путей. Вентральная область покрышки широко вовлечена в системы вознаграждения, а точнее сама по себе является скоплением множества нервных путей. Вентральная область покрышки также вовлечена в формирование зависимостей от таких веществ как никотин, кокаин, героин, амфетамин.

Орексины или гипокретины — название двух нейропептидов, независимо обнаруженных двумя группами исследователей в 1998 году. Два орексина, Орексин А и Орексин Б, имеют 50-процентную гомологию последовательности, не имея при этом значительной гомологии с остальными пептидами в организме. Орексины синтезируются сравнительно небольшой популяцией клеток латерального гипоталамуса, чьи аксоны достигают практически всех регионов мозга. Считается, что недостаток орексинов в мозге приводит к развитию нарколепсии. Орексины демонстрируют сильно сохранившуюся последовательность аминокислот и найдены у всех позвоночных, что указывает на их раннее появление в эволюции позвоночных.

Эпиталамус — это самая дорсальная (задняя) часть таламического мозга, или, иначе говоря, таламической области — той части промежуточного мозга, куда, помимо эпиталамуса, входят также таламус, субталамус и метаталамус, но не входят гипоталамус и гипофиз, причисляемые к гипоталамической области. Эпиталамус возвышается над таламусом. В число его структур входят поводок эпиталамуса, также называемый поводком мозга, треугольник поводка, спайка поводков, подспаечный орган и шишковидная железа, а также связывающие их с другими структурами мозга нервные пути, такие, как мозговая полоска таламуса, поводково-межножковый тракт.
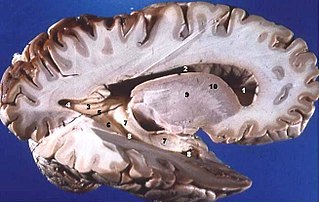
Серое вещество — главный компонент центральной нервной системы позвоночных животных и человека, включающий клеточные тела нейронов, нейропиль, глиальные клетки, а также капилляры. Противопоставляется белому веществу мозга, не содержащему тел нейронов и состоящему главным образом из пучков миелиновых волокон. Цветовая дифференциация белого и серого вещества нервной ткани обусловлена белым цветом миелина. Серое вещество живых тканей имеет серо-коричневую окраску, которую придают кровеносные капилляры и клеточные тела нейронов.

Сосцевидные тела — два маленьких округлых образования, лежащие впереди от заднего продырявленного вещества и сзади от серого бугра, в основании гипоталамуса непосредственно за гипофизом.

Красное ядро — структура в среднем мозге, участвующая в координации движений. Оно состоит из хвостовой магноцеллюлярной (крупноклеточной) и ростральной — парвоцеллюлярной (мелкоклеточной) части. Красное ядро находится в покрышке среднего мозга рядом с чёрной субстанцией. Красное ядро и чёрная субстанция являются подкорковыми центрами экстрапирамидной двигательной системы.

Паравентрикулярное ядро гипоталамуса — группа нейронов гипоталамуса. PVN образуют 3 группы крупноклеточных и 5 групп мелкоклеточных нейронов, выделенных на основе характеристик цитоархитектоники и биохимических свойств. Несмотря на существенные различия в указанных параметрах между группами, каждая группа содержит нейроны, идущие в гипофиз, и нейроны, содержащие вазопрессин и/или окситоцин.
Крупноклеточные нейроны — представлены передней, задней и медиальной обособленными группами, погруженными в матрикс, образованный мелкоклеточными нейронами. Большая часть клеток отдает проекции в гипофиз.
Мелкоклеточные нейроны — образуют матрикс, разделенный на перивентрикулярный, передний, задний, медиальный и латеральные отделы. Основная часть нейронов отдают проекции в ствол мозга.

Субталамус, иначе называемый преталамус или периталамус, вентральный таламус — это часть таламического мозга, или, иначе говоря, таламической области — той части промежуточного мозга, куда, помимо субталамуса, входят также таламус, эпиталамус и метаталамус, но не входят гипоталамус и гипофиз, относимые к гипоталамической области. Его наиболее выраженной структурой является так называемое субталамическое ядро. Субталамус соединён нервными связями с бледным шаром, базальным ядром в конечном мозге.

Передние ядра таламуса, или передняя группа ядер таламуса, — это совокупность таламических ядер, находящихся в ростральной части дорсального таламуса. К передней группе ядер таламуса причисляют антеромедиальное ядро, антеродорсальное ядро, антеровентральное ядро. К передней группе ядер таламуса нередко относят также латеральное дорсальное ядро таламуса.
Магноцеллюлярные клетки гипоталамуса — крупные нейроэндокринные клетки, расположенные в паравентрикулярных и супраоптических ядрах гипоталамуса. Также их можно найти в небольших количествах между указанными выше ядрами. Аксоны магноцеллюлярных клеток идут от гипоталамуса в заднюю долю гипофиза, образуя гипоталамо-гипофизарный тракт, состоящий из паравентрикуло-гипофизарного и супраоптико-гипофизарного пучков. Различают два типа магноцеллюлярных клеток: вазопрессин-продуцирующие и окситоцин-продуцирующие, но есть некоторое количество клеток, которые могут продуцировать оба гормона. Клетки нейровозбудимы и реагируют на афферентную стимуляцию. Большая часть окончаний афферентных нейронов находится на дендритах. Строение этих клеток хорошо изучено на крысах. Магноцеллюлярные клетки имеют один аксон, выходящий в задний гипофиз и образующий около 10 000 нейросекреторных терминалей. Аксон имеет набухания по всей длине, в которых находится множество везикул с гормоном. Эти везикулы выбрасывают гормон экзоцитозом при прохождении потенциала действия по аксону. Кроме аксона, клетка несёт 2—3 дендрита, также содержащих везикулы с гормоном. Таким образом, окситоцин и вазопрессин могут выделяться через терминали аксона в кровь в задней доле гипофиза и через дендриты в мозг. Однако высвобождение гормонов из дендритов не сопровождается постоянными периферическими выбросами, поскольку регуляция секреции в аксоне и дендрите разная. Выброс в дендрите может быть вызван деполяризацией или выбросом внутренних запасов ионов кальция. Вероятно, гормон выделяется в мозг также и через аксоны. Либо аксоны некоторой субпопуляции клеток проецируются в мозг, либо аксоны части клеток дают коллатерали.
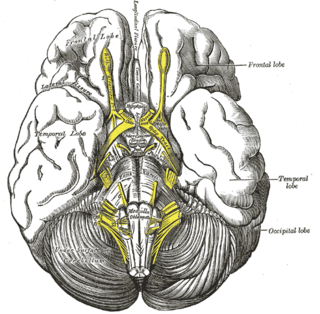
Межножковая ямка — приблизительно ромбовидной формы заполненная спинномозговой жидкостью полость в основании мозга, омывающая снизу ликвором ряд базальных структур головного мозга. Спереди межножковая ямка ограничена перекрёстом зрительных нервов, сзади — передне-верхней поверхностью варолиева моста, с передне-боковых поверхностей — постепенно сходящимися впереди в перекрёсте зрительных нервов зрительными трактами, а с задне-боковых поверхностей — постепенно расходящимися ножками мозга.

Вентролатеральное преоптическое ядро — небольшой участок нейронов в переднем ядре гипоталамуса, расположенный над перекрёстом зрительных нервов в головном мозге человека и животных. Один из центров сна головного мозга. Вместе с восходящей активизирующей ретикулярной системой и системой орексинных аксонов латерального гипоталамуса образует связанную систему нейронов, контролирующих состояния бодрствования, сна, а также переходы между двумя этими состояниями. Вентролатеральное преоптическое ядро активизируется в процессе сна, и выделяет тормозные нейромедиаторы, в основном ГАМК и галанин, которые ингибируют нейроны восходящей активизирующей ретикулярной формации, отвечающие за поддержание бодрствования. Активизация вентролатерального преоптического ядра осуществляется нейромедиаторами серотонином и аденозином, а также простагландином D2. Ингибирование вентролатерального преоптического ядра осуществляется нейромедиаторами норадреналином и ацетилхолином. Изучение данной области и её значение в регулировании сна и бодрствования, включая нарушения сна, является предметом повышенного интереса у нейробиологов.

Супраоптическое ядро гипоталамуса — группа крупноклеточных нейронов в медиальной части гипоталамуса, насчитывающая около 3000 клеток. Важнейшей функцией нейронов супраоптического ядра является синтез вазопрессина, или антидиуретического гормона (АДГ), который путем аксонного транспорта поступает и депонируется в нейрогипофизе. Здесь аксоны нейронов супраоптического ядра образуют синаптоподобные контакты с кровеносными сосудами гипофиза. Вазопрессин накапливается в пресинаптических везикулах и секретируется в кровь при возбуждении нейрона.
 Головной мозг на срединном продольном разрезе.
Головной мозг на срединном продольном разрезе. Свод и мозолистое тело снизу.
Свод и мозолистое тело снизу.