Симбиогенез
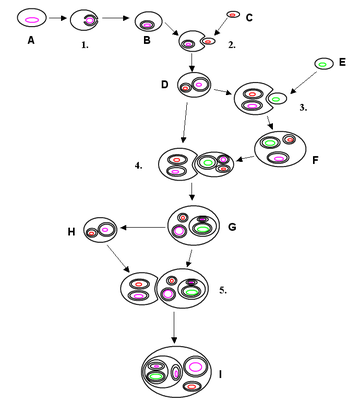
1 — образование двойной мембраны ядра,
2 — приобретение митохондрий,
3 — приобретение пластид,
4 — внедрение получившейся фотосинтезирующей эукариотической клетки в нефотосинтезирующую (например, в ходе эволюции криптофитовых водорослей),
5 — внедрение получившейся клетки снова в нефотосинтезирующую (например, при симбиозе этих водорослей с инфузориями).
Цветом обозначен геном
предков эукариот, митохондрий и пластид.
Теория симбиогене́за (симбиотическая теория, эндосимбиотическая теория, теория эндосимбиоза) объясняет механизм возникновения некоторых органоидов эукариотической клетки — митохондрий, гидрогеносом и пластид.
История
Теорию эндосимбиотического происхождения хлоропластов впервые предложил в 1883 году Андреас Шимпер[1], показавший их саморепликацию внутри клетки. Её возникновению предшествовал вывод А. С. Фаминцына[2] и О. В. Баранецкого о двойственной природе лишайников — симбиотического комплекса гриба и водоросли (1867 год). К. С. Мережковский[3] в 1905 году предложил само название «симбиогенез», впервые детально сформулировал теорию и даже создал на её основе новую систему органического мира. Фаминцын в 1907 году, опираясь на работы Шимпера, также пришёл к выводу, что хлоропласты являются симбионтами, как и водоросли в составе лишайников.
В 1920-е годы теория была развита Б. М. Козо-Полянским, было высказано предположение, что симбионтами являются и митохондрии. Затем долгое время о симбиогенезе практически не упоминали в научной литературе. Второе рождение расширенная и конкретизированная теория получила уже в работах Линн Маргулис начиная с 1960-х годов.
В результате изучения последовательности оснований в митохондриальной ДНК были получены весьма убедительные доводы в пользу того, что митохондрии — это потомки аэробных бактерий (прокариот), родственных риккетсиям, поселившихся некогда в предковой эукариотической клетке и «научившимися» жить в ней в качестве симбионтов (организмов, участвующих в симбиозе). Теперь митохондрии есть почти во всех эукариотических клетках, размножаться вне клетки они уже не способны.
Существуют свидетельства того, что первоначально эндосимбиотические предки митохондрий не могли ни импортировать белки, ни экспортировать АТФ[4]. Вероятно, первоначально они получали от клетки-хозяина пируват, а выгода для хозяина состояла в обезвреживании аэробными симбионтами токсичного для нуклеоцитоплазмы кислорода.
Пластиды, подобно митохондриям, имеют свои собственные прокариотические ДНК и рибосомы. По-видимому, хлоропласты произошли от фотосинтезирующих бактерий, поселившихся в своё время в гетеротрофных клетках протистов, превратив их в автотрофные водоросли.
Доказательства
Митохондрии и пластиды:
- имеют две полностью замкнутые мембраны. При этом внешняя сходна с мембранами вакуолей, внутренняя — бактерий.
- размножаются бинарным делением (причём иногда делятся независимо от деления клетки) и не образуются de novo, то есть не путем синтеза из других органоидов, как, например, лизосома, образующаяся из комплекса Гольджи, а он, в свою очередь, из ЭПС
- генетический материал — кольцевая ДНК, не связанная с гистонами (По доле ГЦ ДНК митохондрий и пластид ближе к ДНК бактерий, чем к ядерной ДНК эукариот)
- имеют свой аппарат синтеза белка — рибосомы и др.
- рибосомы прокариотического типа — c константой седиментации 70S. По строению 16s рРНК близки к бактериальной.
- некоторые белки этих органелл похожи по своей первичной структуре на аналогичные белки бактерий и не похожи на соответствующие белки цитоплазмы.
Проблемы
- ДНК митохондрий и пластид, в отличие от ДНК большинства прокариот, содержат интроны.
- В собственной ДНК митохондрий и хлоропластов закодирована только часть их белков, а остальные закодированы в ДНК ядра клетки. В ходе эволюции происходило «перетекание» части генетического материала из генома митохондрий и хлоропластов в ядерный геном. Этим объясняется тот факт, что ни хлоропласты, ни митохондрии не могут более существовать (размножаться) независимо.
- Не решён вопрос о происхождении ядерно-цитоплазматического компонента (ЯЦК), захватившего прото-митохондрии. Ни бактерии, ни археи не способны к фагоцитозу, питаясь исключительно осмотрофно. Молекулярно-биологические и биохимические исследования указывают на химерную архейно-бактериальную сущность ЯЦК. Как произошло слияние организмов из двух доменов, также не ясно.
Примеры эндосимбиозов

В наши дни существует ряд организмов, содержащих внутри своих клеток другие клетки в качестве эндосимбионтов. Они, однако, не являются сохранившимися до наших дней первичными эукариотами, у которых симбионты ещё не интегрировались в единое целое и не потеряли своей индивидуальности. Тем не менее они наглядно и убедительно показывают возможность симбиогенеза.
- Mixotricha paradoxa — наиболее интересный с этой точки зрения организм. Для движения она использует более 250 000 бактерий Treponema spirochetes, прикреплённых к поверхности её клетки. Митохондрии у этого организма вторично потеряны, но внутри его клетки есть сферические аэробные бактерии, заменяющие эти органеллы.
- Амёбы рода Pelomyxa также не содержат митохондрий и образуют симбиоз с бактериями.
- Инфузории рода Paramecium постоянно содержат внутри клетки водорослей, в частности, Paramecium bursaria образует эндосимбиоз с зелёными водорослями рода хлорелла (Chlorella).
- Одноклеточная жгутиковая водоросль Cyanophora paradoxa содержит цианеллы — органоиды, напоминающие типичные хлоропласты красных водорослей, но отличающиеся от них наличием тонкой клеточной стенки, содержащей пептидогликан (размер генома цианелл такой же, как у типичных хлоропластов, и во много раз меньше, чем у цианобактерий).
Гипотезы эндосимбиотического происхождения других органелл
Эндосимбиоз — наиболее широко признанная версия происхождения митохондрий и пластид. Но попытки объяснить подобным образом происхождение других органелл и структур клетки не находят достаточных доказательств и наталкиваются на обоснованную критику.
Клеточное ядро, нуклеоцитоплазма
Смешение у эукариот многих свойств, характерных для архей и бактерий, позволило предположить симбиотическое происхождение ядра от метаногенной архебактерии, внедрившейся в клетку миксобактерии. Гистоны, к примеру, обнаружены у эукариот и некоторых архей, кодирующие их гены весьма схожи. Другая гипотеза, объясняющая сочетание у эукариот молекулярных признаков архей и эубактерий, состоит в том, что на некотором этапе эволюции похожие на архей предки нуклеоцитоплазматического компонента эукариот приобрели способность к усиленному обмену генами с эубактериями путём горизонтального переноса генов[5].
В последнее десятилетие сформировалась также гипотеза вирусного эукариогенеза (англ. viral eukaryogenesis). В её основании лежит ряд сходств устройства генетического аппарата эукариот и вирусов: линейное строение ДНК, её тесное взаимодействие с белками и др. Было показано сходство ДНК-полимеразы эукариот и поксивирусов, что сделало именно их предков основными кандидатами на роль ядра[6][7].
Жгутики и реснички
Линн Маргулис в книге Symbiosis in Cell Evolution (1981) предположила в том числе происхождение жгутиков и ресничек от симбиотических спирохет. Несмотря на сходство размеров и строения указанных органелл и бактерий и существование Mixotricha paradoxa, использующей спирохет для движения, в жгутиках не было найдено никаких специфически спирохетных белков. Однако известен общий для всех бактерий и архей белок FtsZ, гомологичный тубулину и, возможно, являющийся его предшественником. Жгутики и реснички не обладают такими признаками бактериальных клеток, как замкнутая наружная мембрана, собственный белоксинтезирующий аппарат и способность к делению. Данные о наличии ДНК в базальных тельцах, появившиеся в 1990-е годы, были впоследствии опровергнуты. Увеличение числа базальных телец и гомологичных им центриолей происходит не путём деления, а путём достраивания нового органоида рядом со старым.
Пероксисомы
Кристиан де Дюв обнаружил пероксисомы в 1965 году. Ему же принадлежит предположение, что пероксисомы были первыми эндосимбионтами эукариотической клетки, позволившими ей выживать при нарастающем количестве свободного молекулярного кислорода в земной атмосфере. Пероксисомы, однако, в отличие от митохондрий и пластид, не имеют ни генетического материала, ни аппарата для синтеза белка. Было показано, что эти органеллы формируются в клетке de novo в ЭПР и нет никаких оснований считать их эндосимбионтами[8].
Примечания
- ↑ Schimper A.E.W. Uber die Entwickelung der Chlorophyllkorner und Farbkorper // Bot. Ztschr. Bd. — 1883. — Т. Bot. Ztschr. Bd 41. S. 105—114.. Архивировано 8 февраля 2012 года.
- ↑ Фаминицын А.С. О роли симбиоза в эволюции организмов // Записки Имп. АН. — 1907. — Т. 20, № 3, вып. 8.
- ↑ Мережковский К.С. Терия двух плазм как основа симбиогенезиса, нового учения о происхождении организмов // Уч. зап. Казанского ун-та. — 1909. — Т. 76.
- ↑ Kurland C. G., Andersson S. G. E. Origin and evolution of the mitochondrial proteome (англ.) // Microbilology and Molecular Biology Reviews. — 2000. — Vol. 64, no. 4. — P. 786—820. — doi:10.1128/MMBR.64.4.786-820.2000. — PMID 11104819. Архивировано 19 июля 2018 года.
- ↑ А. В. Марков, А. М. Куликов. Происхождение эвкариот: выводы из анализа белковых гомологий в трёх надцарствах живой природы. Дата обращения: 17 сентября 2009. Архивировано из оригинала 26 мая 2008 года.
- ↑ Takemura Masaharu. Poxviruses and the Origin of the Eukaryotic Nucleus (англ.) // Journal of Molecular Evolution. — 2001. — May (vol. 52, no. 5). — P. 419—425. — ISSN 0022-2844. — doi:10.1007/s002390010171. — PMID 11443345.
- ↑ Villarreal L. P., DeFilippis V. R. A Hypothesis for DNA Viruses as the Origin of Eukaryotic Replication Proteins (англ.) // Journal of Virology. — 2000. — 1 August (vol. 74, no. 15). — P. 7079—7084. — ISSN 0022-538X. — doi:10.1128/jvi.74.15.7079-7084.2000. — PMID 10888648.
- ↑ Gabaldón Toni, Snel Berend, Zimmeren Frank van, Hemrika Wieger, Tabak Henk, Huynen Martijn A. Origin and evolution of the peroxisomal proteome. (англ.) // Biology Direct. — 2006. — Vol. 1, no. 1. — P. 8. — ISSN 1745-6150. — doi:10.1186/1745-6150-1-8. — PMID 16556314.
См. также
- Лишайники
- Протобионты
- Саламандра и водоросли
- Слизни и хлоропласты водорослей
- Тридакна
Литература
- Кулаев И. С. Происхождение эукариотических клеток // Соросовский Образовательный Журнал, 1998, № 5, с. 17-22.
- Подборка статей по проблеме происхождения эукариот
| В библиографических каталогах |
|---|
| Эволюционные процессы |  | |
|---|---|---|
| Факторы эволюции | ||
| Генетика популяций | ||
| Происхождение жизни | ||
| Исторические концепции | ||
| Современные теории | ||
| Эволюция таксонов | ||