Синапс
| Структура типичного химического синапса |
|---|
 Постсинаптическая мембрана Электро- зависимый канал Ca2+ Насос обратного захвата |
| Основные элементы химического синапса: синаптическая щель, везикулы (синаптические пузырьки), нейромедиатор, рецепторы |
Си́напс (греч. σύναψις, от συνάπτειν — соединение, связь) — место контакта между двумя нейронами или между нейроном и получающей сигнал эффекторной клеткой. Служит для передачи нервного импульса между двумя клетками, причём в ходе синаптической передачи амплитуда и частота сигнала могут регулироваться. Передача импульсов осуществляется химическим путём с помощью медиаторов или электрическим путём, посредством прохождения ионов из одной клетки в другую.
Термин был введён в 1897 г. английским физиологом Чарльзом Шеррингтоном. Однако сам Шеррингтон утверждал, что получил идею этого термина в разговоре от физиолога Майкла Фостера[1].
Классификации синапсов
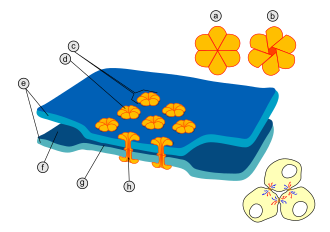
По механизму передачи нервного импульса
- химический — это место близкого прилегания двух нервных клеток, для передачи нервного импульса через которое клетка-источник выпускает в межклеточное пространство особое вещество, нейромедиатор, присутствие которого в синаптической щели возбуждает или затормаживает клетку-приёмник.
- электрический (эфапс) — место более близкого прилегания пары клеток, где их мембраны соединяются с помощью особых белковых образований — коннексонов (каждый коннексон состоит из шести белковых субъединиц). Расстояние между мембранами клетки в электрическом синапсе — 3,5 нм (обычное межклеточное — 20 нм). Так как сопротивление внеклеточной жидкости мало (в данном случае), импульсы через синапс проходят не задерживаясь. Электрические синапсы обычно бывают возбуждающими.
- смешанные синапсы — пресинаптический потенциал действия создает ток, который деполяризует постсинаптическую мембрану типичного химического синапса, где пре- и постсинаптические мембраны неплотно прилегают друг к другу. Таким образом, в этих синапсах химическая передача служит необходимым усиливающим механизмом.
Наиболее распространены химические синапсы. Для нервной системы млекопитающих электрические синапсы менее характерны, чем химические.
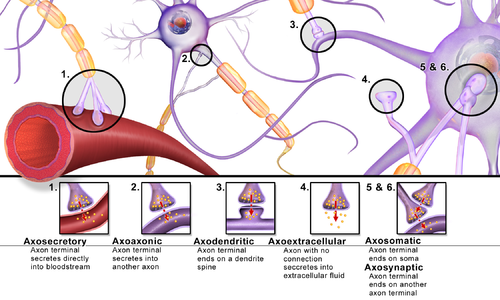
По местоположению и принадлежности структурам
- периферические
- нервно-мышечные
- нейросекреторные (аксо-вазальные)
- рецепторно-нейрональные
- центральные
- аксо-дендритические — с дендритами, в том числе
- аксо-шипиковые — с дендритными шипиками, выростами на дендритах;
- аксо-соматические — с телами нейронов;
- аксо-аксональные — между аксонами;
- дендро-дендритические — между дендритами;
- аксо-дендритические — с дендритами, в том числе
- аминергические, содержащие биогенные амины (например, серотонин, дофамин);
- в том числе адренергические, содержащие адреналин или норадреналин;
- холинергические, содержащие ацетилхолин;
- пуринергические, содержащие пурины;
- пептидергические, содержащие пептиды.
При этом в синапсе не всегда вырабатывается только один медиатор. Обычно основной медиатор выбрасывается вместе с другим, играющим роль модулятора.
По знаку действия синапса
- возбуждающие
- тормозные.
Если первые способствуют возникновению возбуждения в постсинаптической клетке (в них в результате поступления импульса происходит деполяризация мембраны, которая может вызвать потенциал действия при определённых условиях), то вторые, напротив, прекращают или предотвращают его появление, препятствуют дальнейшему распространению импульса. Обычно тормозными являются глицинергические (медиатор — глицин) и ГАМК-ергические синапсы (медиатор — гамма-аминомасляная кислота).
Тормозные синапсы бывают двух видов:
- синапс, в пресинаптических окончаниях которого выделяется медиатор, гиперполяризующий постсинаптическую мембрану и вызывающий возникновение тормозного постсинаптического потенциала;
- аксо-аксональный синапс, обеспечивающий пресинаптическое торможение.
В некоторых синапсах присутствует постсинаптическое уплотнение — электронно-плотная зона, состоящая из белков. По её наличию или отсутствию выделяют синапсы асимметричные и симметричные. Известно, что все глутаматергические синапсы асимметричны, а ГАМКергические — симметричны.
В случаях, когда с постсинаптической мембраной контактирует несколько синаптических расширений, образуются множественные синапсы.
К специальным формам синапсов относятся шипиковые аппараты, в которых с синаптическим расширением контактируют короткие одиночные или множественные выпячивания постсинаптической мембраны дендрита. Шипиковые аппараты значительно увеличивают количество синаптических контактов на нейроне и, следовательно, количество перерабатываемой информации. «Не-шипиковые» синапсы называются «сидячими». Например, сидячими являются все ГАМК-ергические синапсы.
Механизм функционирования химического синапса
Типичный синапс — аксо-дендритический химический. Такой синапс состоит из двух частей: пресинаптической, образованной булавовидно расширенным окончанием аксона передающей клетки и постсинаптической, представленной контактирующим участком плазматической мембраны воспринимающей клетки (в данном случае — участком дендрита).
Между обеими частями имеется синаптическая щель — промежуток шириной 10—50 нм между постсинаптической и пресинаптической мембранами, края которой укреплены межклеточными контактами.
Часть аксолеммы булавовидного расширения, прилежащая к синаптической щели, называется пресинаптической мембраной. Участок цитолеммы воспринимающей клетки, ограничивающий синаптическую щель с противоположной стороны, называется постсинаптической мембраной, в химических синапсах она рельефна и содержит многочисленные рецепторы.
В синаптическом расширении имеются мелкие везикулы, так называемые синаптические пузырьки, содержащие либо медиатор (вещество-посредник в передаче возбуждения), либо фермент, разрушающий этот медиатор. На постсинаптической, а часто и на пресинаптической мембранах присутствуют рецепторы к тому или иному медиатору.
При деполяризации пресинаптической терминали открываются потенциал-чувствительные кальциевые каналы, ионы кальция входят в пресинаптическую терминаль и запускают механизм слияния синаптических пузырьков с мембраной. В результате медиатор выходит в синаптическую щель и присоединяется к белкам-рецепторам постсинаптической мембраны, которые делятся на метаботропные и ионотропные. Первые связаны с G-белком и запускают каскад реакций внутриклеточной передачи сигнала. Вторые связаны с ионными каналами, которые открываются при связывании с ними нейромедиатора, что приводит к изменению мембранного потенциала. Медиатор действует в течение очень короткого времени, после чего разрушается специфическим ферментом. Например, в холинэргических синапсах фермент, разрушающий медиатор в синаптической щели — ацетилхолинэстераза. Одновременно часть медиатора может перемещаться с помощью белков-переносчиков через постсинаптическую мембрану (прямой захват) и в обратном направлении через пресинаптическую мембрану (обратный захват). В ряде случаев медиатор также поглощается соседними клетками нейроглии.
Открыты два механизма высвобождения: с полным слиянием везикулы с плазмалеммой и так называемый «поцеловал и убежал» (англ. kiss-and-run), когда везикула соединяется с мембраной, и из неё в синаптическую щель выходят небольшие молекулы, а крупные остаются в везикуле. Второй механизм, предположительно, быстрее первого, с помощью него происходит синаптическая передача при высоком содержании ионов кальция в синаптической бляшке.
Следствием такой структуры синапса является одностороннее проведение нервного импульса. Существует так называемая синаптическая задержка — время, нужное для передачи нервного импульса. Её длительность составляет около — 0,5 мс.
Так называемый «принцип Дейла» (один нейрон — один медиатор) признан ошибочным. Или, как иногда считают, он уточнён: из одного окончания клетки может выделяться не один, а несколько медиаторов, причём их набор постоянен для данной клетки.
История открытия
- В 1897 году Шеррингтон сформулировал представление о синапсах.
- За исследования нервной системы, в том числе синаптической передачи, в 1906 году Нобелевскую премию получили Гольджи и Рамон-и-Кахаль.
- В 1921 австрийский учёный О. Лёви (О. Loewi) установил химическую природу передачи возбуждения через синапсы и роль в ней ацетилхолина. Получил Нобелевскую премию в 1936 г. совместно с Г. Дейлом (Н. Dale).
- В 1933 советский учёный А. В. Кибяков установил роль адреналина в синаптической передаче.
- 1970 — Б. Кац (В. Katz, Великобритания), У. фон Эйлер (U. v. Euler, Швеция) и Дж. Аксельрод (J. Axelrod, США) получили Нобелевскую премию за открытие роли норадреналина в синаптической передаче.
См. также
Примечания
- ↑ French R. D. Some problems and sources in the foundation of modern physiology in Great Britain // Hist. Sci.. — 1971. — № 10. — С. 28-29.
Ссылки
- Савельев А. В. Методология синаптической самоорганизации и проблема дистальных синапсов нейронов // Журнал проблем эволюции открытых систем. — Казахстан, Алматы, 2006. — Т. 8, № 2. — С. 96—104.
- Экклз Д. К. Физиология синапсов. — М.: Мир, 1966. — 397 с.
| Нейроны (Серое вещество) |
|
|---|---|
| Афферентный нерв/ Сенсорный нейрон |
|
| Эфферентный нерв/ Моторный нейрон |
|
| Синапс | |
| Сенсорный рецептор | |
| Нейроглия | |
| Миелин (Белое вещество) | |
| Соединительная ткань |
|













