Соотношение полов
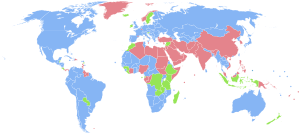
| страны, в которых женщин больше, чем мужчин страны, в которых мужчин больше, чем женщин страны с практически равным соотношением мужчин и женщин нет данных |
Соотношение полов — отношение числа самцов к числу самок в раздельнополой популяции.
Соотношение полов наряду с половым диморфизмом является важной характеристикой раздельнополой популяции. Обычно его выражают количеством людей мужского пола, приходящихся на 100 людей женского пола, долей мужских особей или в процентах. В зависимости от стадии онтогенеза различают первичное, вторичное и третичное соотношение полов. Первичное — это соотношение полов в зиготах после оплодотворения; вторичное — соотношение полов при рождении и, наконец, третичное — соотношение полов зрелых, способных размножаться особей популяции.
Повышенное первичное и вторичное соотношение полов
В настоящее время общепризнано, что у большинства видов животных и растений основным механизмом, определяющим пол, является хромосомный.
Поскольку в ходе гаметогенеза гаметы, содержащие Х- или Y-хромосомы, производятся в равном числе, считалось, что этот механизм обеспечивает примерно равные доли полов при зачатии. Однако, вторичное соотношение полов зависит не только от пропорции гетерогамет, но и от многих других факторов. Например, от отношения скоростей старения и элиминации Х- и Y-несущих спермиев в организме самца, от их способности достичь яйцеклетки и оплодотворить её, от сродства яйцеклеток к Х- или Y-спермиям, наконец, от жизнеспособности мужских и женских эмбрионов на разных стадиях эмбрионального развития.
Давно было замечено, что у многих видов животных вторичное соотношение полов несколько отличается от пропорции 1 : 1 в сторону избытка самцов примерно 105—106 самцов на 100 самок. Больше всего надёжных данных собрано по человеку. Среднее значение вторичного соотношения полов по всем человеческим популяциям составляет около 106. Учёт неодинаковой (дифференциальной) смертности полов во время эмбриональной стадии развития ещё больше сдвигает экстраполированное значение первичного соотношения полов от пропорции 1 : 1. Все доступные данные о половом составе выкидышей и мертворождённых у человека показывают, что плодов мужского пола погибает во время утробной жизни в 2-4 раза больше, чем женского[2]. Таким образом, первичное соотношение полов у человека, по всей видимости, отклоняется от пропорции 1 : 1 в сторону избытка мужских зигот, и оно, видимо, больше, чем вторичное. У человека соотношение полов при рождении может искусственно нарушаться за счёт абортов и детоубийства[3][4][5].
Связь между вторичным и третичным соотношением полов
Между вторичным и третичным соотношением полов существует прямая зависимость — чем больше рождаемость мужских особей, тем больше их может дожить до зрелого возраста.
Многие данные свидетельствуют также о том, что вторичное соотношение полов зависит от третичного. У нескольких видов растений и животных (дрёма, гуппи, клещи (3 вида), дрозофила, мыши, крысы, лесной североамериканский сурок и человек) прямыми экспериментами было показано, что увеличение третичного соотношения полов приводит к уменьшению вторичного[6].
Насекомые
У некоторых насекомых (пчёл и других перепончатокрылых, червецов, клещей) из оплодотворённых яиц получаются самки (или самки и самцы), а из неоплодотворённых развиваются только самцы. Поэтому, чем меньше самцов в исходной популяции, тем в среднем меньше яиц подвергается оплодотворению и тем больше самцов получается в потомстве[7].



Человек
Половой дисбаланс
Гендерный дисбаланс — демографический эффект, возникающий в связи с войнами, например, в Европе после Первой мировой войны и после Второй мировой войны (в России, см. Население СССР), или как результат внутренней политики (напр., одна семья — один ребёнок).
Люди не являются строго моногамными, и у тех народов, у которых распространено многожёнство, возникают различные отклонения третичного соотношения полов от пропорции 1 : 1, как правило в сторону его уменьшения. У нигерийцев антрополог Норткот У. Томас заметил возрастание вторичного соотношения полов с увеличением числа жён[8].
Вторичное соотношение полов в гаремах увеличивается примерно до 62 % (гаремы: императора Чжу Юаньчжана (1328—1398, Китай) 26 мальчиков : 16 девочек; фараона Рамзеса II (1317—1251 д.н. э., Египет) 74 жены, 111:68; и султана Мауля Исмаила (1646—1727, Марокко) 548:340)[9][10][11]. К данным, полученным для стран, где больше ценится рождение мальчиков, следует относиться с осторожностью, принимая во внимание возможность искусственного вмешательства до и после рождения.
Географические различия
В таблице приведены показатели количества мужчин на одну женщину в различных странах по данным Всемирной книги фактов ЦРУ на 2014 год[12].
| Государство/регион | Новорождённые | 0-14 лет | 15-24 лет | 25-54 лет | 55-64 лет | 65 и более | Всё население |
|---|---|---|---|---|---|---|---|
| Афганистан | 1,05 | 1,03 | 1,04 | 1,04 | 1,03 | 0,87 | 1,03 |
| Албания | 1,11 | 1,12 | 1,05 | 0,91 | 0,98 | 0,89 | 0,98 |
| Алжир | 1,05 | 1,05 | 1,05 | 1,02 | 1,03 | 0,84 | 1,03 |
| Американское Самоа | 1,06 | 0,96 | 0,98 | 1,06 | 1,00 | 0,86 | 1,01 |
| Андора | 1,07 | 1,05 | 1,08 | 1,06 | 1,07 | 1,00 | 1,07 |
| Ангола | 1,05 | 1,04 | 1,04 | 1,02 | 1,02 | 0,86 | 1,02 |
| Ангилья (Брит.) | 1,03 | 1,05 | 0,98 | 0,82 | 0,92 | 0,96 | 0,93 |
| Антигуа и Барбуда | 1,05 | 1,03 | 0,98 | 0,84 | 0,90 | 0,76 | 0,90 |
| Аргентина | 1,05 | 1,05 | 1,04 | 1,00 | 0,97 | 0,70 | 0,97 |
| Армения | 1,14 | 1,15 | 1,03 | 0,92 | 0,93 | 0,59 | 0,89 |
| Аруба | 1,02 | 1,01 | 1,01 | 0,93 | 0,90 | 0,65 | 0,90 |
| Австралия | 1,06 | 1,05 | 1,05 | 1,03 | 1,01 | 0,85 | 1,01 |
| Австрия | 1,05 | 1,05 | 1,04 | 1,01 | 0,95 | 0,73 | 0,95 |
| Азербайджан | 1,12 | 1,16 | 1,07 | 0,95 | 0,98 | 0,62 | 0,98 |
| Багамские острова | 1,03 | 1,03 | 1,03 | 1,00 | 0,96 | 0,62 | 0,96 |
| Бахрейн | 1,03 | 1,03 | 1,28 | 1,90 | 1,54 | 0,91 | 1,54 |
| Бангладеш | 1,04 | 1,03 | 0,88 | 0,90 | 0,95 | 0,96 | 0,95 |
| Барбадос | 1,01 | 1,00 | 1,00 | 0,99 | 0,94 | 0,65 | 0,94 |
| Белоруссия | 1,06 | 1,06 | 1,06 | 0,96 | 0,87 | 0,46 | 0,87 |
| Бельгия | 1,05 | 1,04 | 1,04 | 1,02 | 0,96 | 0,72 | 0,96 |
| Белиз | 1,05 | 1,04 | 1,04 | 1,02 | 1,03 | 0,90 | 1,03 |
| Бенин | 1,05 | 1,04 | 1,04 | 1,02 | 1,01 | 0,67 | 1,01 |
| Бермудские о-ва (Брит.) | 1,02 | 1,02 | 1,00 | 1,00 | 0,94 | 0,72 | 0,94 |
| Бутан | 1,05 | 1,04 | 1,04 | 1,15 | 1,09 | 1,11 | 1,10 |
| Боливия | 1,05 | 1,04 | 1,03 | 0,95 | 0,98 | 0,79 | 0,98 |
| Босния и Герцеговина | 1,07 | 1,07 | 1,07 | 1,01 | 0,95 | 0,63 | 0,95 |
| Ботсвана | 1,03 | 1,04 | 0,99 | 1,12 | 1,03 | 0,67 | 1,02 |
| Бразилия | 1,05 | 1,04 | 1,03 | 0,98 | 0,97 | 0,74 | 0,98 |
| Виргинские о-ва (Брит.) | 1,05 | 0,97 | 0,91 | 0,90 | 0,92 | 0,95 | 0,93 |
| Бруней | 1,05 | 1,07 | 0,98 | 0,94 | 0,99 | 0,95 | 1,00 |
| Болгария | 1,06 | 1,05 | 1,06 | 0,99 | 0,92 | 0,68 | 0,92 |
| Буркина-Фасо | 1,03 | 1,00 | 1,01 | 1,03 | 0,99 | 0,62 | 0,99 |
| Мьянма | 1,06 | 1,04 | 1,03 | 0,99 | 0,99 | 0,77 | 0,99 |
| Бурунди | 1,03 | 1,01 | 1,00 | 1,00 | 0,99 | 0,67 | 0,98 |
| Кабо-Верде | 1,03 | 1,01 | 1,00 | 0,94 | 0,94 | 0,60 | 0,94 |
| Камбоджа | 1,05 | 1,02 | 0,98 | 0,95 | 0,94 | 0,60 | 0,94 |
| Камерун | 1,03 | 1,02 | 1,01 | 1,01 | 1,01 | 0,84 | 1,01 |
| Канада | 1,06 | 1,05 | 1,06 | 1,03 | 0,99 | 0,79 | 0,99 |
| Каймановы о-ва (Брит.) | 1,02 | 1,01 | 0,98 | 0,95 | 0,95 | 0,90 | 0,95 |
| Центральноафриканская Республика | 1,03 | 1,01 | 1,01 | 1,00 | 0,98 | 0,66 | 0,98 |
| Чад | 1,04 | 1,03 | 0,94 | 0,83 | 0,93 | 0,72 | 0,93 |
| Чили | 1,04 | 1,04 | 1,04 | 0,99 | 0,97 | 0,71 | 0,97 |
| Китай | 1,11 | 1,16 | 1,13 | 1,05 | 1,06 | 0,92 | 1,06 |
| Колумбия | 1,06 | 1,05 | 1,04 | 0,98 | 0,98 | 0,72 | 0,98 |
| Коморские о-ва | 1,03 | 0,99 | 0,94 | 0,90 | 0,94 | 0,92 | 0,94 |
| Демократическая Республика Конго | 1,03 | 1,02 | 1,01 | 1,00 | 0,99 | 0,72 | 0,99 |
| Республика Конго | 1,03 | 1,02 | 0,99 | 1,03 | 1,01 | 0,69 | 0,99 |
| Острова Кука (Нов. Зел.) | 1,04 | 1,13 | 1,15 | 1,01 | 1,07 | 0,96 | 1,07 |
| Коста-Рика | 1,05 | 1,05 | 1,04 | 1,01 | 1,01 | 0,86 | 1,01 |
| Кот-д’Ивуар | 1,03 | 1,02 | 1,02 | 1,05 | 1,02 | 0,96 | 1,03 |
| Хорватия | 1,06 | 1,06 | 1,05 | 0,98 | 0,93 | 0,66 | 0,93 |
| Куба | 1,06 | 1,06 | 1,05 | 1,01 | 0,99 | 0,82 | 0,99 |
| Кюрасао (Нид.) | 1,15 | 1,05 | 1,08 | 0,92 | 0,79 | 0,71 | 0,92 |
| Кипр | 1,05 | 1,06 | 1,19 | 1,10 | 1,04 | 0,77 | 1,04 |
| Чехия | 1,06 | 1,06 | 1,05 | 1,05 | 0,97 | 0,67 | 0,95 |
| Дания | 1,06 | 1,05 | 1,04 | 1,00 | 0,97 | 0,80 | 0,97 |
| Джибути | 1,03 | 1,01 | 0,89 | 0,71 | 0,85 | 0,82 | 0,86 |
| Доминика | 1,05 | 1,05 | 1,06 | 1,03 | 1,02 | 0,77 | 1,02 |
| Доминиканская Республика | 1,04 | 1,04 | 1,04 | 1,05 | 1,03 | 0,86 | 1,03 |
| Эквадор | 1,05 | 1,04 | 1,03 | 0,95 | 0,99 | 0,92 | 0,99 |
| Египет | 1,05 | 1,05 | 1,05 | 1,03 | 1,02 | 0,82 | 1,03 |
| Сальвадор | 1,05 | 1,05 | 1,01 | 0,85 | 0,93 | 0,80 | 0,93 |
| Экваториальная Гвинея | 1,03 | 1,03 | 1,04 | 0,98 | 0,99 | 0,75 | 0,99 |
| Эритрея | 1,03 | 1,01 | 1,00 | 0,98 | 0,98 | 0,79 | 0,98 |
| Эстония | 1,06 | 1,06 | 1,06 | 0,92 | 0,84 | 0,49 | 0,84 |
| Эфиопия | 1,03 | 1,00 | 0,99 | 0,99 | 0,99 | 0,83 | 0,99 |
| Европейский Союз | 1,06 | 1,05 | 1,05 | 1,02 | 0,94 | 0,74 | 0,96 |
| Фарерские о-ва (Дат.) | 1,07 | 1,08 | 1,07 | 1,18 | 1,09 | 0,93 | 1,09 |
| Фиджи | 1,05 | 1,05 | 1,05 | 1,05 | 1,03 | 0,85 | 1,03 |
| Финляндия | 1,04 | 1,04 | 1,04 | 1,04 | 0,96 | 0,71 | 0,96 |
| Франция | 1,05 | 1,05 | 1,05 | 1,01 | 0,96 | 0,74 | 0,96 |
| Французская Полинезия | 1,05 | 1,06 | 1,07 | 1,05 | 1,05 | 0,95 | 1,05 |
| Габон | 1,03 | 1,01 | 1,00 | 1,00 | 0,99 | 0,73 | 0,99 |
| Гамбия | 1,03 | 1,01 | 0,98 | 0,96 | 0,98 | 0,90 | 0,98 |
| Сектор Газа | 1,06 | 1,06 | 1,05 | 1,05 | 1,04 | 0,68 | 1,04 |
| Грузия | 1,08 | 1,12 | 1,08 | 0,94 | 0,92 | 0,66 | 0,91 |
| Германия | 1,06 | 1,06 | 1,04 | 1,03 | 0,97 | 0,76 | 0,97 |
| Гана | 1,03 | 1,01 | 0,99 | 0,94 | 0,97 | 0,88 | 0,98 |
| Гибралтар (Брит.) | 1,07 | 1,05 | 1,09 | 1,01 | 1,01 | 0,95 | 1,01 |
| Греция | 1,06 | 1,06 | 1,04 | 0,99 | 0,96 | 0,78 | 0,96 |
| Гренландия (Дат.) | 1,05 | 1,03 | 1,03 | 1,12 | 1,10 | 1,10 | 1,11 |
| Гренада | 1,10 | 1,07 | 0,99 | 1,05 | 1,03 | 0,83 | 1,02 |
| Гуам (США) | 1,06 | 1,06 | 1,07 | 1,04 | 1,03 | 0,84 | 1,03 |
| Гватемала | 1,05 | 1,04 | 1,01 | 0,91 | 0,97 | 0,87 | 0,97 |
| Гернси (Брит.) | 1,05 | 1,07 | 1,04 | 1,01 | 0,98 | 0,81 | 0,98 |
| Гвинея | 1,03 | 1,02 | 1,02 | 1,01 | 1,00 | 0,79 | 1,00 |
| Гвинея-Бисау | 1,03 | 1,00 | 0,99 | 0,99 | 0,95 | 0,64 | 0,95 |
| Гайана | 1,05 | 1,04 | 1,06 | 1,09 | 1,02 | 0,69 | 0,99 |
| Гайти | 1,01 | 1,01 | 1,00 | 0,99 | 0,99 | 0,80 | 0,98 |
| Гондурас | 1,05 | 1,04 | 1,04 | 1,02 | 1,01 | 0,79 | 1,01 |
| Сянган (Гонконг) (Кит.) | 1,13 | 1,14 | 1,05 | 0,75 | 0,87 | 0,88 | 0,94 |
| Венгрия | 1,06 | 1,06 | 1,06 | 1,01 | 0,91 | 0,59 | 0,91 |
| Исландия | 1,04 | 1,03 | 1,02 | 1,02 | 1,00 | 0,85 | 1,00 |
| Индия | 1,12 | 1,13 | 1,13 | 1,06 | 1,08 | 0,91 | 1,08 |
| Индонезия | 1,05 | 1,04 | 1,04 | 1,03 | 1,00 | 0,78 | 1,00 |
| Иран | 1,05 | 1,05 | 1,06 | 1,04 | 1,03 | 0,89 | 1,03 |
| Ирак | 1,05 | 1,04 | 1,03 | 1,04 | 1,03 | 0,87 | 1,03 |
| Ирландия | 1,06 | 1,04 | 1,04 | 1,01 | 1,00 | 0,84 | 1,00 |
| Мэн (Брит.) | 1,08 | 1,11 | 1,08 | 0,99 | 1,00 | 0,85 | 0,99 |
| Израиль | 1,05 | 1,05 | 1,05 | 1,05 | 1,01 | 0,78 | 1,01 |
| Италия | 1,06 | 1,05 | 1,01 | 0,98 | 0,93 | 0,74 | 0,93 |
| Ямайка | 1,05 | 1,04 | 1,01 | 0,97 | 0,98 | 0,81 | 0,98 |
| Япония | 1,06 | 1,07 | 1,09 | 0,98 | 0,94 | 0,76 | 0,95 |
| Джерси (Брит.) | 1,06 | 1,07 | 1,04 | 1,00 | 0,97 | 0,74 | 0,96 |
| Иордания | 1,06 | 1,05 | 1,05 | 1,01 | 1,02 | 0,95 | 1,03 |
| Казахстан | 0,94 | 1,00 | 1,04 | 0,95 | 0,92 | 0,52 | 0,92 |
| Кения | 1,02 | 1,01 | 1,00 | 1,02 | 1,00 | 0,79 | 1,00 |
| Кирибати | 1,05 | 1,04 | 1,00 | 0,93 | 0,96 | 0,64 | 0,96 |
| КНДР | 1,05 | 1,03 | 1,02 | 1,00 | 0,94 | 0,51 | 0,94 |
| Республика Корея | 1,07 | 1,08 | 1,13 | 1,04 | 1,00 | 0,69 | 1,00 |
| Косово | 1,08 | 1,08 | 1,11 | 1,12 | 1,06 | 0,73 | 1,06 |
| Кувейт | 1,05 | 1,08 | 1,22 | 1,74 | 1,42 | 0,96 | 1,43 |
| Киргизия | 1,07 | 1,05 | 1,03 | 0,96 | 0,96 | 0,63 | 0,96 |
| Лаос | 1,04 | 1,02 | 0,99 | 0,97 | 0,99 | 0,82 | 0,99 |
| Латвия | 1,05 | 1,05 | 1,04 | 0,99 | 0,86 | 0,48 | 0,86 |
| Ливан | 1,05 | 1,05 | 1,04 | 1,02 | 1,00 | 0,86 | 0,96 |
| Лесото | 1,03 | 1,01 | 0,91 | 0,96 | 0,98 | 0,99 | 0,97 |
| Либерия | 1,03 | 1,02 | 0,95 | 1,00 | 0,99 | 1,01 | 1,00 |
| Ливия | 1,05 | 1,05 | 1,07 | 1,10 | 1,08 | 1,04 | 1,08 |
| Лихтенштейн | 1,26 | 1,14 | 0,98 | 1,00 | 0,99 | 0,81 | 0,98 |
| Литва | 1,06 | 1,05 | 1,05 | 1,00 | 0,89 | 0,53 | 0,89 |
| Люксембург | 1,07 | 1,06 | 1,05 | 1,00 | 0,97 | 0,71 | 0,97 |
| Аомынь (Макао) (Кит.) | 1,05 | 1,10 | 1,08 | 0,81 | 0,91 | 0,88 | 0,91 |
| Македония | 1,08 | 1,07 | 1,07 | 1,03 | 0,99 | 0,75 | 0,99 |
| Мадагаскар | 1,03 | 1,02 | 1,00 | 1,00 | 1,00 | 0,83 | 1,00 |
| Малави | 1,02 | 0,99 | 0,99 | 0,98 | 0,98 | 0,75 | 0,99 |
| Малайзия | 1,07 | 1,06 | 1,03 | 1,03 | 1,03 | 0,89 | 1,03 |
| Мальдивская республика | 1,05 | 1,04 | 1,40 | 1,43 | 1,29 | 0,94 | 1,34 |
| Мали | 1,03 | 1,01 | 0,91 | 0,87 | 0,95 | 1,01 | 0,95 |
| Мальта | 1,06 | 1,05 | 1,06 | 1,04 | 0,99 | 0,79 | 0,99 |
| Маршалловы о-ва | 1,05 | 1,04 | 1,04 | 1,04 | 1,04 | 0,95 | 1,04 |
| Мавритания | 1,03 | 1,01 | 0,96 | 0,86 | 0,93 | 0,74 | 0,93 |
| Маврикий | 1,05 | 1,04 | 1,02 | 1,00 | 0,97 | 0,67 | 0,97 |
| Мексика | 1,05 | 1,05 | 1,02 | 0,93 | 0,96 | 0,81 | 0,96 |
| Федеративные Штаты Микронезии | 1,05 | 1,03 | 1,01 | 0,94 | 0,98 | 0,77 | 0,99 |
| Молдавия | 1,06 | 1,07 | 1,07 | 1,00 | 0,95 | 0,60 | 0,94 |
| Монако | 1,04 | 1,05 | 1,05 | 0,99 | 0,95 | 0,81 | 0,95 |
| Монголия | 1,05 | 1,04 | 1,02 | 0,94 | 0,96 | 0,77 | 1,00 |
| Черногория | 1,07 | 0,95 | 0,89 | 1,17 | 0,99 | 0,66 | 0,99 |
| Монтсеррат (Брит.) | 1,04 | 1,06 | 1,11 | 0,91 | 1,00 | 1,55 | 1,00 |
| Марокко | 1,05 | 1,03 | 0,99 | 0,94 | 0,97 | 0,82 | 0,97 |
| Мозамбик | 1,02 | 1,01 | 0,95 | 0,88 | 0,95 | 0,85 | 0,95 |
| Намибия | 1,03 | 1,02 | 1,02 | 1,09 | 1,02 | 0,78 | 1,02 |
| Науру | 0,84 | 0,79 | 1,07 | 0,99 | 0,90 | 0,78 | 0,91 |
| Непал | 1,04 | 1,04 | 1,01 | 0,91 | 0,97 | 0,87 | 0,96 |
| Нидерланды | 1,05 | 1,05 | 1,04 | 1,01 | 0,98 | 0,80 | 0,98 |
| Новая Каледония (Фр.) | 1,05 | 1,05 | 1,04 | 1,01 | 1,00 | 0,81 | 1,00 |
| Новая Зеландия | 1,05 | 1,05 | 1,05 | 1,00 | 0,99 | 0,85 | 0,99 |
| Никарагуа | 1,05 | 1,04 | 1,01 | 0,90 | 0,96 | 0,83 | 0,96 |
| Нигер | 1,03 | 1,02 | 0,98 | 1,00 | 1,01 | 1,03 | 1,01 |
| Нигерия | 1,06 | 1,05 | 1,05 | 1,05 | 1,04 | 0,85 | 1,01 |
| Северные Марианские о-ва (США) | 1,06 | 1,09 | 1,23 | 0,73 | 0,93 | 0,91 | 0,93 |
| Норвегия | 1,06 | 1,05 | 1,06 | 1,06 | 1,01 | 0,78 | 0,98 |
| Оман | 1,05 | 1,05 | 1,10 | 1,41 | 1,20 | 1,03 | 1,22 |
| Пакистан | 1,05 | 1,06 | 1,07 | 1,08 | 1,06 | 0,89 | 1,06 |
| Палау | 1,05 | 1,07 | 0,99 | 1,56 | 1,10 | 0,40 | 1,12 |
| Панама | 1,05 | 1,04 | 1,04 | 1,03 | 1,01 | 0,86 | 1,02 |
| Папуа-Новая Гвинея | 1,05 | 1,04 | 1,03 | 1,07 | 1,05 | 1,14 | 1,05 |
| Парагвай | 1,05 | 1,04 | 1,01 | 1,00 | 1,01 | 0,87 | 1,01 |
| Перу | 1,05 | 1,04 | 1,00 | 0,93 | 0,97 | 0,90 | 0,97 |
| Филиппины | 1,05 | 1,04 | 1,04 | 1,01 | 1,00 | 0,76 | 1,00 |
| Польша | 1,06 | 1,06 | 1,04 | 1,01 | 0,94 | 0,62 | 0,94 |
| Португалия | 1,07 | 1,09 | 1,13 | 1,02 | 0,95 | 0,69 | 0,95 |
| Пуэрто-Рико (США) | 1,02 | 1,05 | 1,04 | 0,91 | 0,92 | 0,77 | 0,92 |
| Катар | 1,02 | 1,03 | 2,75 | 4,80 | 3,37 | 1,50 | 3,29 |
| Румыния | 1,06 | 1,06 | 1,05 | 1,02 | 0,95 | 0,68 | 0,95 |
| Россия | 1,06 | 1,06 | 1,05 | 0,96 | 0,86 | 0,44 | 0,86 |
| Руанда | 1,03 | 1,02 | 1,00 | 1,01 | 0,99 | 0,67 | 0,99 |
| Сен-Бартелеми (Фр.) | 1,03 | 1,05 | 1,11 | 1,20 | 1,13 | 1,00 | 1,14 |
| Святой Елены, Вознесения и Тристан-да-Кунья, о-ва (Брит.) | 1,05 | 1,04 | 1,04 | 0,98 | 1,02 | 1,01 | 1,02 |
| Сент-Китс и Невис | 1,02 | 1,00 | 0,97 | 1,04 | 1,00 | 0,80 | 1,00 |
| Сент-Люсия | 1,06 | 1,06 | 1,03 | 0,93 | 0,95 | 0,83 | 0,95 |
| Сен-Мартен (Фр.) | 1,04 | 0,99 | 1,03 | 0,90 | 0,93 | 0,81 | 0,93 |
| Сен-Пьер и Микелон (Фр.) | 1,10 | 1,07 | 1,06 | 0,99 | 0,96 | 0,69 | 0,97 |
| Сент-Винсент и Гренадины | 1,03 | 1,02 | 1,02 | 1,08 | 1,03 | 0,84 | 1,03 |
| Самоа | 1,05 | 1,07 | 1,05 | 1,08 | 1,05 | 0,78 | 1,05 |
| Сан-Марино | 1,10 | 1,14 | 1,05 | 0,89 | 0,94 | 0,80 | 0,94 |
| Сан-Томе и Принсипи | 1,03 | 1,04 | 1,03 | 0,96 | 1,00 | 0,84 | 1,00 |
| Саудовская Аравия | 1,05 | 1,05 | 1,15 | 1,33 | 1,20 | 1,08 | 1,21 |
| Сенегал | 1,03 | 1,01 | 0,99 | 0,84 | 0,94 | 0,85 | 0,94 |
| Сербия | 1,07 | 1,07 | 1,06 | 1,02 | 0,95 | 0,69 | 0,95 |
| Сейшельские о-ва | 1,03 | 1,05 | 1,10 | 1,11 | 1,05 | 0,61 | 1,04 |
| Сьерра-Леоне | 1,03 | 0,99 | 0,94 | 0,92 | 0,94 | 0,78 | 0,94 |
| Сингапур | 1,07 | 1,05 | 0,97 | 0,95 | 0,96 | 0,82 | 0,96 |
| Синт-Мартен (Нид.) | 1,05 | 1,08 | 0,96 | 0,95 | 0,94 | 0,92 | 0,97 |
| Словакия | 1,07 | 1,05 | 1,06 | 1,02 | 0,94 | 0,60 | 0,94 |
| Словения | 1,07 | 1,06 | 1,05 | 1,02 | 0,95 | 0,66 | 0,95 |
| Соломоновы о-ва | 1,05 | 1,06 | 1,06 | 1,04 | 1,04 | 0,95 | 1,04 |
| Сомали | 1,03 | 1,00 | 1,03 | 1,07 | 1,01 | 0,66 | 1,01 |
| ЮАР | 1,02 | 1,01 | 1,01 | 1,07 | 0,98 | 0,67 | 0,99 |
| Испания | 1,07 | 1,06 | 1,07 | 1,04 | 0,98 | 0,74 | 0,97 |
| Шри-Ланка | 1,04 | 1,04 | 1,03 | 0,96 | 0,96 | 0,75 | 0,96 |
| Судан | 1,05 | 1,03 | 1,07 | 0,94 | 1,02 | 1,24 | 1,02 |
| Суринам | 1,05 | 1,04 | 1,04 | 1,04 | 1,01 | 0,77 | 1,01 |
| Свазиленд | 1,03 | 1,02 | 1,02 | 1,06 | 1,00 | 0,69 | 0,99 |
| Швеция | 1,06 | 1,06 | 1,06 | 1,04 | 1,00 | 0,81 | 0,98 |
| Швейцария | 1,06 | 1,06 | 1,04 | 1,01 | 0,97 | 0,76 | 0,97 |
| Сирия | 1,06 | 1,05 | 1,03 | 1,00 | 1,01 | 0,85 | 1,03 |
| Китайская Республика (Тайвань) | 1,07 | 1,07 | 1,06 | 1,00 | 0,99 | 0,89 | 1,00 |
| Таджикистан | 1,05 | 1,04 | 1,03 | 0,98 | 0,99 | 0,73 | 0,99 |
| Танзания | 1,03 | 1,02 | 1,00 | 1,01 | 0,99 | 0,76 | 0,99 |
| Таиланд | 1,05 | 1,05 | 1,04 | 0,97 | 0,97 | 0,82 | 0,98 |
| Восточный Тимор | 1,07 | 1,06 | 1,03 | 0,93 | 1,01 | 0,96 | 1,01 |
| Того | 1,03 | 1,01 | 1,00 | 0,99 | 0,98 | 0,77 | 0,98 |
| Тонга | 1,03 | 1,03 | 1,05 | 1,00 | 1,01 | 0,86 | 1,01 |
| Тринидад и Тобаго | 1,03 | 1,04 | 1,08 | 1,08 | 1,03 | 0,76 | 1,03 |
| Тунис | 1,07 | 1,07 | 1,01 | 0,95 | 0,99 | 0,96 | 0,99 |
| Турция | 1,05 | 1,05 | 1,04 | 1,02 | 1,02 | 0,84 | 1,02 |
| Туркменистан | 1,05 | 1,03 | 1,01 | 0,98 | 0,98 | 0,77 | 0,98 |
| Тёркс и Кайкос, о-ва (Брит.) | 1,05 | 1,04 | 0,96 | 1,03 | 1,02 | 0,80 | 1,02 |
| Тувалу | 1,05 | 1,05 | 1,12 | 0,95 | 0,97 | 0,73 | 0,97 |
| Уганда | 1,03 | 0,99 | 0,99 | 1,00 | 0,99 | 0,80 | 0,99 |
| Украина | 1,07 | 1,06 | 1,04 | 0,94 | 0,85 | 0,49 | 0,85 |
| Объединённые Арабские Эмираты | 1,05 | 1,05 | 1,47 | 3,22 | 2,19 | 1,77 | 2,19 |
| Великобритания | 1,05 | 1,05 | 1,04 | 1,04 | 0,99 | 0,80 | 0,99 |
| США | 1,05 | 1,05 | 1,05 | 1,00 | 0,97 | 0,77 | 0,97 |
| Уругвай | 1,04 | 1,04 | 1,03 | 0,97 | 0,93 | 0,66 | 0,93 |
| Узбекистан | 1,06 | 1,05 | 1,03 | 0,99 | 0,99 | 0,75 | 0,99 |
| Вануату | 1,05 | 1,04 | 0,99 | 0,96 | 1,00 | 1,07 | 1,01 |
| Венесуэла | 1,05 | 1,04 | 1,01 | 0,97 | 0,98 | 0,79 | 0,98 |
| Вьетнам | 1,12 | 1,11 | 1,07 | 1,01 | 1,00 | 0,62 | 1,00 |
| Виргинские о-ва (США) | 1,06 | 1,02 | 0,85 | 0,83 | 0,87 | 0,83 | 0,88 |
| Уоллис и Футуна (Фр.) | 1,05 | 1,09 | 1,10 | 0,99 | 1,03 | 0,89 | 1,02 |
| Западный берег реки Иордан | 1,06 | 1,05 | 1,05 | 1,06 | 1,04 | 0,71 | 1,04 |
| Западная Сахара | 1,04 | 1,02 | 1,01 | 0,97 | 0,99 | 0,78 | 0,98 |
| Мир | 1,07 | 1,07 | 1,062 | 1,022 | 0,955 | 0,802 | 1,014 |
| Йемен | 1,05 | 1,04 | 1,03 | 1,04 | 1,03 | 0,90 | 1,03 |
| Замбия | 1,03 | 1,01 | 1,00 | 1,02 | 1,00 | 0,75 | 1,00 |
| Зимбабве | 1,03 | 1,02 | 1,01 | 1,07 | 1,00 | 0,70 | 0,95 |
В России
В конце XIX века на 1000 мужчин приходилось 1055 женщин, а в 20-30-е годы XX века — около 1120. После Великой Отечественной войны половой дисбаланс в России резко увеличился[13]. Число женщин на 1000 мужчин: в 1959—1242; в 1970—1193; в 1979—1174; в 1989—1140[14].
| Экономический район | Численность мужчин (тыс.чел) | Численность женщин (тыс.чел) | Количество женщин на 1000 мужчин |
|---|---|---|---|
| Северо-западный | 3982 | 4759 | 1195 |
| Центрально-Чернозёмный | 3441 | 4076 | 1184 |
| Волго-Вятский | 3652 | 4306 | 1179 |
| Центральный | 14105 | 16377 | 1161 |
| Поволжский | 7645 | 8875 | 1160 |
| Уральский | 9187 | 10596 | 1153 |
| Северо-Кавказский | 8874 | 10036 | 1131 |
| Западно-Сибирский | 6957 | 7835 | 1126 |
| Восточно-Сибирский | 4032 | 4504 | 1117 |
| Северный | 2475 | 2760 | 1115 |
| Дальневосточный | 3255 | 3438 | 1056 |
| Число женщин на 1 мужчину | Число женщин на 1 мужчину | Число женщин на 1 мужчину | Число женщин на 1 мужчину | Число мужчин на 1 женщину | Число мужчин на 1 женщину | Число мужчин на 1 женщину | Число мужчин на 1 женщину | |
|---|---|---|---|---|---|---|---|---|
| При структуре 1989 и соотношении полов | При структуре 2002 и соотношении полов | При структуре 1989 и соотношении полов | При структуре 2002 и соотношении полов | При структуре 1989 и соотношении полов | При структуре 2002 и соотношении полов | При структуре 1989 и соотношении полов | При структуре 2002 и соотношении полов | |
| 1989 | 1989 | 2002 | 2002 | 1989 | 1989 | 2002 | 2002 | |
| 0-4 | 0,086 | 0,047 | 0,085 | 0,046 | 0,078 | 0,042 | 0,079 | 0,042 |
| 5-9 | 0,081 | 0,051 | 0,08 | 0,05 | 0,074 | 0,045 | 0,075 | 0,046 |
| 10-14 | 0,076 | 0,076 | 0,075 | 0,075 | 0,069 | 0,068 | 0,07 | 0,069 |
| 15-19 | 0,071 | 0,091 | 0,072 | 0,093 | 0,065 | 0,086 | 0,064 | 0,084 |
| 20-24 | 0,07 | 0,083 | 0,071 | 0,084 | 0,063 | 0,076 | 0,062 | 0,075 |
| 25-29 | 0,09 | 0,076 | 0,092 | 0,078 | 0,081 | 0,07 | 0,079 | 0,069 |
| 30-34 | 0,093 | 0,072 | 0,094 | 0,073 | 0,083 | 0,064 | 0,081 | 0,063 |
| 35-39 | 0,085 | 0,075 | 0,088 | 0,077 | 0,074 | 0,066 | 0,072 | 0,065 |
| 40-44 | 0,057 | 0,093 | 0,058 | 0,096 | 0,048 | 0,081 | 0,047 | 0,078 |
| 45-49 | 0,061 | 0,09 | 0,061 | 0,09 | 0,048 | 0,071 | 0,048 | 0,071 |
| 50-54 | 0,075 | 0,079 | 0,076 | 0,08 | 0,057 | 0,061 | 0,056 | 0,06 |
| 55-59 | 0,068 | 0,044 | 0,068 | 0,044 | 0,048 | 0,031 | 0,047 | 0,031 |
| 60-64 | 0,075 | 0,076 | 0,069 | 0,07 | 0,041 | 0,039 | 0,045 | 0,042 |
| 65-69 | 0,046 | 0,083 | 0,032 | 0,058 | 0,017 | 0,021 | 0,025 | 0,032 |
| 70-74 | 0,038 | 0,079 | 0,028 | 0,057 | 0,013 | 0,019 | 0,018 | 0,026 |
| 75-79 | 0,037 | 0,047 | 0,033 | 0,043 | 0,01 | 0,012 | 0,012 | 0,013 |
| 80-84 | 0,02 | 0,019 | 0,02 | 0,018 | 0,005 | 0,004 | 0,005 | 0,004 |
| 85+ | 0,011 | 0,014 | 0,01 | 0,013 | 0,002 | 0,002 | 0,002 | 0,002 |
| Все возрасты | 1,139 | 1,195 | 1,112 | 1,146 | 0,877 | 0,858 | 0,887 | 0,871 |
| Все возрасты по переписи населения | 1,140 | 1,140 | 0,877 | 0,872 |
«Феномен военных лет»
Значительный дефицит мужского пола появляется во время и после войн. В этот период в воюющих странах наблюдается увеличение мужских рождений примерно на 1—2 %. Так, во время Первой мировой войны в европейских странах доля мальчиков возросла на 1-2,5 % по сравнению с обычной и составила в Германии 108,5 %, примерно такое же увеличение произошло к концу 1942 года в Великобритании и Франции. В Закавказье с начала 90-х годов (конфликт Армении и Азербайджана по поводу Нагорного Карабаха, грузино-абхазский конфликт) отмечается рост рождаемости мальчиков. Это явление, установленное на очень большом статистическом материале, получило в демографии название «феномена военных лет», поскольку соотношение полов у человека в мирные годы довольно стабильно[2][16][17][18].
Убедительного объяснения роста доли рождений мальчиков во время и после войн наука пока не дала; были выдвинуты различные гипотезы[19][20][21].
Понижение третичного соотношения полов в онтогенезе
В ходе онтогенеза соотношение полов у многих видов растений, животных и человека понижается. Это связано с повышенной смертностью и повреждаемостью мужских систем по сравнению с соответствующими женскими. Эта картина наблюдается почти на всех стадиях онтогенеза и на всех уровнях организации, исследуем ли мы различные виды (человек, животные или растения), разные уровни организации (особь, орган, ткань или клетка) или устойчивость к разным вредным факторам среды (низкие и высокие температуры, голод, яды, паразиты, болезни и др.)[22][23][24][25][26].
Гамильтон (Hamilton, 1948) приводит обзор дифференциальной смертности полов для 70 видов, включая такие различные формы жизни, как нематоды, моллюски, ракообразные, насекомые, паукообразные, птицы, рептилии, рыбы и млекопитающие. Согласно этим данным, у 62 видов (89 %) средняя длительность жизни самцов короче, чем у самок; у большинства остальных нет разницы, и лишь в отдельных случаях продолжительность жизни самцов больше, чем у самок[27].
Можно заключить, что повышенная смертность мужского пола — общебиологическое явление, оно наблюдается у растений, животных и человека для всех уровней организации от всех экстремальных значений факторов среды.
У человека средняя продолжительность жизни различна в разных странах, и у мужчин на 3-10 лет меньше, чем у женщин.
Повышенная смертность особей мужского пола, как общебиологическое явление, может быть объяснена у некоторых организмов неслучайными причинами. Например, у многих млекопитающих, включая оленей, человека и других животных, возможна повышенная смертность самцов в процессе борьбы самцов за право обладания самками. У некоторых паукообразных, у которых самки гораздо крупнее самцов, наблюдается систематическое поедание самками самцов после спаривания. У пчёл все рабочие пчелы являются недоразвитыми самками, не способными давать потомства. Настоящая самка пчёл — матка в улье всегда одна. Если их оказывается две, то одна из них погибает. Настоящие самцы пчёл — трутни после спаривания с маткой обратно в улей не пускаются и вскоре погибают.
Влияние генетических факторов на вторичное соотношение полов
Определение пола ребёнка у человека не является чисто случайным событием. Исследованиями, проведёнными в Саксонии в 1876—1885 гг. и позже в Англии, Франции, США и Финляндии, было установлено, что семьи, в которых превалирует один пол, появляются значительно чаще, а семьи, с равным соотношением полов — гораздо реже по сравнению с теоретическим ожиданием. Было также установлено, что имеется слабая, но реальная тенденция в семьях производить второго ребёнка того же пола, что и первый, третьих детей одинакового пола со вторыми и т. д[2].
У крупного рогатого скота в потомстве отдельных быков производителей наблюдали преобладание бычков (1,5 %), а также корреляцию между соотношением полов в потомстве производителя и его отца[28].
Влияние репродуктивного ранга («успеха») на вторичное соотношение полов
Репродуктивный ранг (доступ к брачным партнёрам) родителей влияет на соотношение полов их потомства. У самцов репродуктивный ранг коррелирует, как правило, с их социально-иерархическим рангом. У самок может иметь место обратная корреляция, поскольку их иерархический ранг, как и у самцов, определяется силой и агрессивностью, а репродуктивный ранг — больше привлекательностью и уступчивостью. У оленей, свиней, овец, собак, тюленей и человека было показано, что самки, имеющие, больший «репродуктивный успех», рожали больше потомков мужского пола[29].
Влияние факторов среды на вторичное соотношение полов
Количество пыльцы и вторичное соотношение полов у растений
Было обнаружено, что количество пыльцы, попадающее на женский цветок может влиять на вторичное соотношение полов у перекрёстноопыляющихся растений. Эта зависимость была подтверждена у четырёх видов растений, принадлежащих к трём семействам — щавель (Rumex acetosa, Polygonaceae),[30][31] дрёма (Melandrium album, Cariophyllaceae),[32][33] конопля (Cannabis sativa, Cannabinaceae)[34] и хмель (Humulus japonicus, Cannabinaceae)[35].
Температура яиц
У многих видов черепах, ящериц, крокодилов и некоторых змей пол потомства зависит от температуры инкубации яиц. Так как самки определяют место откладывания яиц, они могут контролировать пол потомства. Температура при которой достигается соотношение полов 1 : 1 называется пороговой температурой. У некоторых видов черепах низкие температуры инкубации приводят к появлению самцов, а высокие — самок, у других видов самцы появляются как при низких, так и при высоких температурах, а самки — при средних.
Вторичное соотношение полов и интенсивность половой деятельности
Интенсивность (частота) половой деятельности у животных может влиять на вторичное соотношение полов. Она непосредственно связана с физиологическими параметрами организма. Низкая интенсивность половой деятельности у самцов и самок равносильна тому, что в процессе оплодотворения участвуют, как правило, более старые спермии и яйцеклетки. У семи видов животных принадлежащих к шести семействам (кур, мышей, кроликов, свиней, лошадей, баранов и крупного рогатого скота) при уменьшении половой деятельности самцов или старении спермы наблюдалось уменьшение числа рождений мужских особей[6].
Для человека увеличение рождаемости мальчиков при повышении интенсивности половой деятельности мужчин доказано на большом статистическом материале в работах Джеймса[36].
Различия в инактивации и гибели Y- и X-спермиев
Связь вторичного соотношения полов с интенсивностью половой деятельности у человека и разных видов животных может быть обусловлена более быстрой гибелью или инактивацией Y-спермиев по сравнению с X-спермиями[37]. Для человека это предположение было подтверждено экспериментально. Было обнаружено, что после длительных периодов воздержания содержание Y-хроматина в сперме заметно уменьшается. При воздержании менее 2 дней процент Y-хроматина составлял 43,5 %, при воздержании 14 дней и более — 37,2 %. Уменьшением концентрации Y-сперматозоидов в сперме с течением времени можно объяснить заниженные по сравнению с теоретически ожидаемой величиной (50 %) значения Y-хроматина, полученные разными авторами, а также большой разброс этих значений[38].
Старение яйцеклеток
Многими авторами была отмечена связь между задержкой оплодотворения яйцеклеток и вторичным соотношением полов. В классических работах Гертвига и Кушакевича на лягушке (Rana esculenta) было отмечено повышение рождения мужских особей[39][40]. Сходные результаты были получены на бабочках, шелкопряде, форели, клопах, дрозофиле, мышах, крысах, кроликах, крупном рогатом скоте и человеке, всего 15 видов как с мужской, так и с женской гетерогаметностью, принадлежащих к 11 семействам[6].
Зависимость вторичного соотношения полов от возраста матери
В. Н. Большаков и Б. С. Кубанцев проанализировав зависимость соотношения полов потомства от возраста матери у лисиц, норок, песцов, собак, свиней, овец, крупного рогатого скота, лошадей и человека пришли к выводу, что «…у молодых матерей в потомстве, как правило, преобладают новорожденные мужского пола. У средневозрастных матерей, находящихся в расцвете своей воспроизводительной функции, возрастает относительное число потомков женского пола. У матерей старшей возрастной группы вновь увеличивается в потомстве процент особей мужского пола.»[41]
Зависимость вторичного соотношения полов от веса матери
Есть данные, что женщины с небольшим весом, а также женщины, набравшие существенный вес за время беременности чаще рожают девочек[42].
Зависимость вторичного соотношения полов от стресса
Есть данные, что как мужчины, так и женщины, подверженные хроническим стрессам, например работающие водителями, пилотами или дайверами, чаще рождают девочек[42].
Зависимость вторичного соотношения полов от условий жизни
Для целых стран, чем лучше условия жизни, благосостояние, климат, питание, тем меньше рождается мальчиков. Такую же закономерность отмечают и животноводы — чем лучше условия содержания сельскохозяйственных животных, тем больше рождается самок[43].
Во время больших природных или социальных сдвигов (резкие изменения климата, засуха, война, голод, переселение) наблюдается тенденция повышения вторичного соотношения полов — растёт процент мальчиков[2][16][17].
Однако, есть ряд ученых, которые доказывают прямо противоположное, что, наоборот, в более благоприятных условиях чаще рождаются именно мальчики[44].
Современная медицина позволяет определять пол пренатально[англ.] . Это используется в некоторых странах для выбора пола потомства[англ.], преимущественно путём абортирования плодов нежелательного пола, также при зачатии или после рождения (детоубийство)[45]. Подобная практика при широком использовании может приводить даже к нарушению поло-возрастной структуры общества, чем объясняется аномально большое количество лиц мужского пола в возрастной категории до 30 лет в современных Китае и Индии. (См. также Биоэтика).
Эволюционные механизмы, регулирующие соотношение полов в популяции
Начало работам по эволюции соотношения полов было положено «принципом Фишера»[46]. Согласно этой теории, для видов, выращивающих своё потомство, третичное соотношение полов 1 : 1 является оптимальным и обеспечивает наилучшие условия для выживания потомства. Чтобы объяснить наблюдаемые у многих видов отклонения от этого соотношения, Фишер утверждал, что равным должно быть не число самцов и самок, а затраты родителей на производство потомства мужского или женского пола до завершения воспитания потомства, то есть чем «дороже обходятся» родителям потомки данного пола, тем их меньше производится. Из «теории равных затрат» Фишера вытекает, в частности, что в тех случаях, когда потомки разного пола имеют разные размеры, должно наблюдаться нарушение соотношения полов. Однако данные Хоува (Howe, 1977) по вороньим дроздам (Quiscalus quiscula), у которых самцы тяжелее самок на 20 %, и большое исследование Ньютона и Маркисса (Newton, Marquiss, 1978) по ястребу (Accipiter nisus), у которого, наоборот, самки в 2 раза массивнее самцов, не подтвердили предсказания теории Фишера[47][48]. Гамильтон (Hamilton, 1967) обратил внимание на ошибочность теории Фишера во всех случаях, когда имеет место локальная конкуренция за скрещивания. Он насчитал около 25 видов клещей и насекомых из 16 разных семейств, у которых постоянный значительный инбридинг сочетается с большим избытком самок и с арренотокической системой размножения[49].
По мнению Калмуса и Смита (Kalmus, Smith, 1960), значение третичного соотношения полов 1 : 1 оптимально, поскольку максимально облегчает встречу особей противоположного пола и снижает степень инбридинга[50]. Их теория не в состоянии объяснить наблюдаемые отклонения значений вторичного соотношения полов у многих видов от 1 : 1, а также его изменения в зависимости от различных факторов.
Ближе всех к идее регуляции соотношения полов подошёл Мэйнард Смит (1981), который предположил, что «родителям может быть выгодно производить потомство пола, более редкого в данной местности»[51].
Организменные механизмы регуляции соотношения полов
Связующим звеном обратной связи у перекрестноопыляемых растений служит количество пыльцы, попадающее на женский цветок, а у животных — интенсивность половой деятельности, которая проявляется через неодинаковое старение Х- и Y-спермиев и через разное сродство к ним свежих и старых яйцеклеток. При этом малые количества пыльцы, интенсивная половая деятельность самцов, свежая сперма и старые яйцеклетки — те факторы, которые приводят к увеличению рождаемости мужских особей.
Популяционные механизмы регуляции соотношения полов
Для реализации популяционного механизма необходимо, чтобы вероятность иметь потомка данного пола различалась у разных особей и определялась их генотипом. При этом должна существовать обратная зависимость между репродуктивным рангом данной особи и полом её потомства: чем выше репродуктивный ранг, тем больше должно быть потомков противоположного пола. В этом случае регуляция может осуществляться на популяционном уровне, большим или меньшим участием в размножении особей, дающих в потомстве избыток самцов или самок.
Существование отрицательной обратной связи показано прямыми экспериментами по крайней мере у трёх видов (у одного вида растений и двух видов животных): Melandrium album, Lebistes reticulatus peters, Macrocheles. Многие виды растений и животных (4 вида растений и 16 видов животных), а также человек обладают теми или иными механизмами для её реализации[6].
См. также
Примечания
- ↑ Data from the CIA World Factbook [1] Архивная копия от 12 августа 2008 на Wayback Machine. Map compiled in 2021, data from 2020.
- ↑ 1 2 3 4 Штерн К. Основы генетики человека. — М.: Медицина, 1965.
- ↑ Coale, Ansley J.; Banister, Judith. Five decades of missing females in China (неопр.) // Demography. — Springer-Verlag, 1994. — Т. 31, № 3. — С. 459—479. — doi:10.2307/2061752. — PMID 7828766. Архивировано 24 марта 2018 года.
- ↑ Mungello, D. E. The Great Encounter of China and the West, 1500–1800 (англ.). — Rowman & Littlefield, 2012. — ISBN 9781442219755. Архивировано 4 апреля 2015 года.
- ↑ Mashru, Ram (2012-01-18). "It's a girl: The three deadliest words in the world". The Independent. Архивировано 12 декабря 2013.
- ↑ 1 2 3 4 Геодакян В. А., Геодакян С. В. (1958) Существует ли отрицательная обратная связь в определении пола? Журнал общей биологии. 46 № 2, с. 201—216.
- ↑ Flanders S. E. (1946) Control of sex and sex-limited polymorphism in the Hymenoptera.—Quart. Rev. Biol. 21 № 2, p. 135—143.
- ↑ Northcote Whitridge Thomas (1868—1936) Report on the Ibo-speaking peoples of Nigeria, Pt. 1. L. (1913), Rart II. — Demography
- ↑ У Хань. Жизнеописание Чжу Юаньчжана. М.: Наука, 1980.
- ↑ Эберс Г. Уарда. М.: Наука, 1965.
- ↑ Азия и Африка сегодня, 1970, № 5, с. 53—54.
- ↑ Sex ratio Архивная копия от 16 октября 2013 на Wayback Machine [] // Всемирная книга фактов ЦРУ
- ↑ Женщин в России на 10 миллионов больше, чем мужчин // Демоскоп Weekly : сайт. — 24 октября - 6 ноября 2005. — № 219—220. Архивировано 21 марта 2018 года.
- ↑ 1 2 Евгений Андреев Анатолий Вишневский Екатерина Кваша Татьяна Харькова. Российская половозрастная пирамида. Диспропорция полов в России продолжает ослабевать // Демоскоп Weekly : сайт. — 26 сентября - 9 октября 2005. — № 215—216. Архивировано 4 февраля 2018 года.
- ↑ Е.Е. Лейзерович. Различия в соотношении числа мужчин и числа женщин на территории России (статистический обзор) = Опубликовано в книге: Трансформация российского пространства: социально-экономические и природно-ресурсные факторы (полимасштабный анализ). Сборник докладов XXV сессии МАРС / ред. С.С. Артоболевский, Л.М. Синцеров. М.: ИГ РАН, 2008. с. 173-181 // Демоскоп Weekly : сайт. — 8 - 21 марта 2010. — № 413—414. Архивировано 31 января 2018 года.
- ↑ 1 2 Новосельский С. А. (1958) Вопросы демографической и санитарной статистики. М., Госстатиздат, с. 1-197.
- ↑ 1 2 Scheinfield A. (1944) Women and men. New York, 405 p.
- ↑ Heath C. W. (1954) Physique, temperament and sex ratio. Hum. Biol.. 26 N 4, p. 337—342.
- ↑ James, William H (March 2009). "The variations of human sex ratio at birth during and after wars, and their potential explanations". Journal of Theoretical Biology. 257 (1): 116—23. Bibcode:2009JThBi.257..116J. doi:10.1016/j.jtbi.2008.09.028. PMID 18952111. Архивировано 23 мая 2022. Дата обращения: 26 декабря 2020.
- ↑ Allen, Laura Why Does War Breed More Boys? (амер. англ.). Popular Science (18 декабря 2008). Дата обращения: 12 июля 2023. Архивировано 12 июля 2023 года.
- ↑ С.А. Новосельский. Влияние войны на половой состав рождающихся//Вопросы демографической и санитарной статистики (Избранные произведения) / под ред. А.М. Меркова Москва, "Медгиз", 1958 с. 191-199. Дата обращения: 5 февраля 2024. Архивировано 5 февраля 2024 года.
- ↑ Левин В. Л. (1949) К вопросу о различной повреждаемости тканей самцов и самок. Опыты с белыми мышами. Докл. AН СССР. 66 № 4. с. 749—751.
- ↑ Левин В. Л. (1951) Некоторые особенности действия стрихнина на изолированные ткани и целые организмы самцов и самок грызунов. Докл. AН СССР. 78 № 1, с. 165—168.
- ↑ Левин В. Л. (1951) К вопросу о различной повреждаемости тканей самцов и самок. Опыты с белыми крысами. Докл. AН СССР. 78 № 4. с. 817—819.
- ↑ Лэк Д. (1957) Численность животных и её регуляция в природе. М., Изд-во иностр. лит. 400 с.
- ↑ Светлов П. Г., Светлова М. Г. (1950) Происхождение половых различий повреждаемости в онтогенезе двудомных цветковых растений. Докл. AН СССР. 70 № 5. с. 925—928.
- ↑ Hamilton J. B. (1948) The role of testicular secretions as indicated by the effects of castration in man and by studies of pathological conditions and the short life span associated with maleness. Recent Progress in Hormone Research, v. 3, N.Y., Acad. Press, p. 257—322.
- ↑ Bar-Anon R., Robertson A. (1975) Variation in sex ratio between progeny groups in dairy cattle. Theoret. Appl. Genet. 46 p. 63-65.
- ↑ Trivers R. L., Willard D. E. (1973) Natural selection of parental ability to vary the sex ratio of offspring. Science. 179 № 4068, p. 90—92.
- ↑ Соrrens С. (1922) Geschlechtsbestimmung und Zahlenverhaltnis der Geschlechter beim Sauerampfer (Rumex acetosa). «Biol. Zbl.» 42, 465—480.
- ↑ Rychlewski J., Kazlmierez Z. (1975) Sex ratio in seeds of Rumex acetosa L. as a result of sparse or abundant pollination. «Acta Biol. Cracov» Scr. Bot., 18, 101—114.
- ↑ Correns C. (1928) Bestimmung, Vererbung und Verteilung des Geschlechter bei den hoheren Pflanzen. Handb. Vererbungswiss., 2, 1-138.
- ↑ Mulcahy D. L. (1967) Optimal sex ratio in Silene alba. «Heredity» 22 № 3, 41.
- ↑ Riede W. (1925) Beitrage zum Geschlechts- und Anpassungs-problem. «Flora» 18/19
- ↑ Kihara H., Hirayoshi J. (1932) Die Geschlechtschromosomen von Humulus japonicus. Sieb. et. Zuce. In: 8th Congr. Jap. Ass. Adv. Sci., p. 363—367 (cit.: Plant Breeding Abstr., 1934, 5, № 3, p. 248, ref. № 768).
- ↑ James W. H. (1976) Timing of fertilization and sex ratio of offspring. A review. Ann. Hum. Biol. 3 N 6, p. 549—556.
- ↑ Геодакян В. А., Кособутский В. И. (1967) Регуляция соотношения полов механизмом обратной связи. ДАН СССР. 173 № 4, с. 938—941.
- ↑ Schwinger E., Ites J., Korte B. (1976) Studies on frequency of Y-chromatin in human sperm. Hum. Genet. 34 N 3, p. 265—270.
- ↑ Hertwig R. (1912) Ober den derzeitigen Stand des Sexualitatsproblems.— Biol. Zentralblatt. В. 32.
- ↑ Кушакевич С. (1910) История развития половых желез у Rana esculenta. СПб.
- ↑ Большаков В. Н., Кубанцев Б. С. (1964) Половая структура популяций млекопитающих и её динамика. М., Наука, 233 с.
- ↑ 1 2 Битва зародышей. Дата обращения: 5 февраля 2024. Архивировано 5 февраля 2024 года.
- ↑ Милованов В. К. (1962) Биология воспроизведения и искусственное осеменение животных. М., Сельхозгиз, 696 с.
- ↑ Горветт, Зарайя (2016-11-09). "Почему у миллиардеров чаще рождаются сыновья". BBC Русская служба (англ.). Архивировано 3 августа 2017. Дата обращения: 14 июля 2017.
- ↑ Preventing gender-biased sex selection: an interagency statement Архивная копия от 23 декабря 2014 на Wayback Machine // WHO, 2011
- ↑ Fisher R. A. (1930) The genetical theory of natural selection. Univ. Press, Oxford.
- ↑ Howe H. F. (1977) Sex ratio adjustment in the common grakle. Science. 198 p. 744—746.
- ↑ Newton J., Marquiss M. (1978) Sex ratio among nestlings of the European sparrowhawk. Am. Nat., (cit.: Maynard Smith J., 1978).
- ↑ Hamilton W. D. (1967) Extraordinary sex ratios. Science. 156 p. 477—488.
- ↑ Kalmus H., Smith A. B. (1960) Evolutionary origin of sexual differentiation and the sex-ratio. Nature. 186 N 4730, p. 1004—1006.
- ↑ Мэйнард Смит Дж. (1981) Эволюция полового размножения. М., Мир, 271 с.
| В библиографических каталогах |
|---|










