Глюконеогене́з — метаболический путь, приводящий к образованию глюкозы из неуглеводных соединений. Наряду с гликогенолизом, этот путь поддерживает в крови уровень глюкозы, необходимый для работы многих тканей и органов, в первую очередь, нервной ткани и эритроцитов. Он служит важным источником глюкозы в условиях недостаточного количества гликогена, например, после длительного голодания или тяжёлой физической работы. Глюконеогенез является обязательной частью цикла Кори, кроме того, этот процесс может быть использован для превращения пирувата, образованного при дезаминировании аминокислот аланина и серина.

Ци́кл трикарбо́новых кисло́т (сокр. ЦТК, цикл Кре́бса, цитра́тный цикл, цикл лимо́нной кислоты́) — центральная часть общего пути катаболизма, циклический биохимический процесс, в ходе которого ацетильные остатки (СН3СО-) окисляются до диоксида углерода (CO2). При этом за один цикл образуется 2 молекулы CO2, 3 НАДН, 1 ФАДH2 и 1 ГТФ (или АТФ). Электроны, находящиеся на НАДН и ФАДH2, в дальнейшем переносятся на дыхательную цепь, где в ходе реакций окислительного фосфорилирования образуется АТФ.

γ-Аминомасляная кислота — органическое соединение, непротеиногенная аминокислота, важнейший тормозной нейромедиатор центральной нервной системы (ЦНС) человека и других млекопитающих. Аминомасляная кислота является биогенным веществом. Содержится в ЦНС и принимает участие в нейромедиаторных и метаболических процессах в мозге.
Гетероцисты — дифференцированные клетки нитчатых цианобактерий, осуществляющие азотфиксацию. При недостатке соединений азота в среде они появляются регулярно вдоль трихомы из вегетативных клеток и акинет. Цианобактерии — фототрофы, осуществляющие оксигенный фотосинтез, однако кислород, атмосферный и выделяемый при фотосинтезе, ингибирует фермент нитрогеназу, необходимую для азотфиксации, поэтому у нитчатых цианобактерий в процессе эволюции возникли специализированные клетки для азотфиксации.
Дыхательная цепь переноса электронов, также электрон-транспортная цепь (сокр. ЭТЦ, англ. ETC, Electron transport chain) — система трансмембранных белков и переносчиков электронов, необходимых для поддержания энергетического баланса. ЭТЦ поддерживает баланс за счёт переноса электронов и протонов из НАД∙Н и ФАДН2 в акцептор электронов. В случае аэробного дыхания акцептором может быть молекулярный кислород (О2). В случае анаэробного дыхания акцептором могут быть NO3−, NO2−, Fe3+, фумарат, диметилсульфоксид, сера, SO42−, CO2 и т. д. ЭТЦ у прокариот локализована в ЦПМ, у эукариот — на внутренней мембране митохондрий. Переносчики электронов расположены в порядке уменьшения сродства к электрону, то есть по своему окислительно-восстановительному потенциалу, где у акцептора самое сильное сродство к электрону. Поэтому транспорт электрона на всём протяжении цепи протекает самопроизвольно с выделением энергии. Выделение энергии в межмембранное пространство при переносе электронов происходит ступенчато, в виде протона (H+). Протоны из межмембранного пространства попадают в протонную помпу, где наводят протонный потенциал. Протонный потенциал преобразуется АТФ-синтазой в энергию химических связей АТФ. Сопряжённая работа ЭТЦ и АТФ-синтазы носит название окислительного фосфорилирования.

Окисли́тельное фосфорили́рование — метаболический путь, при котором энергия, образовавшаяся при окислении питательных веществ, запасается в митохондриях клеток в виде АТФ. Хотя различные формы жизни на Земле используют разные питательные вещества, АТФ является универсальным соединением, в котором запасается энергия, необходимая для других метаболических процессов. Почти все аэробные организмы осуществляют окислительное фосфорилирование. Вероятно, широкому распространению этого метаболического пути способствовала его высокая энергетическая эффективность по сравнению с анаэробным брожением.
Декарбоксилирование аминокислот — процесс отщепления карбоксильной группы аминокислот в виде CO2.

Кето́новые тела́ (синоним: ацето́новые тела, ацето́н [распространённый медицинский жаргонизм]) — группа продуктов обмена веществ, которые образуются в печени из ацетил-КоА:
- ацетон (пропанон) H3C−CO−CH3,
- ацетоуксусная кислота (ацетоацетат) H3C−CO−CH2−COOH,
- бета-оксимасляная кислота (β-гидроксибутират) H3C−CHOH−CH2−COOH.

ФАД — флавинадениндинуклеотид — кофермент, принимающий участие во многих окислительно-восстановительных биохимических процессах. ФАД существует в двух формах — окисленной и восстановленной, его биохимическая функция, как правило, заключается в переходе между этими формами.

Карбоксисо́мы (полиэдральные тела) — микрокомпартменты в клетках бактерий, содержащие фиксирующие углерод ферменты. Они представляют собой многогранные однослойные белковые тела полиэдрической формы от 80 до 140 нанометров в диаметре. Они являются основной частью механизма концентрирования CO2, что помогает преодолеть неэффективность рибулозодифосфаткарбоксилазы (Рубиско) — главного фермента, лимитирующего скорость фиксации углерода в цикле Кальвина. Эти органеллы обнаружены во всех цианобактериях и многих хемотрофных бактериях, фиксирующих CO2.
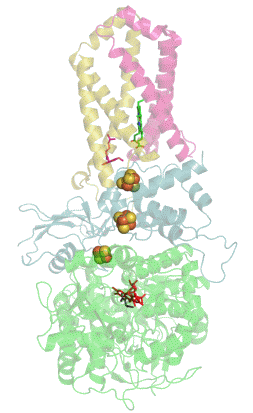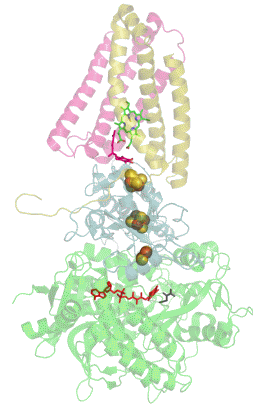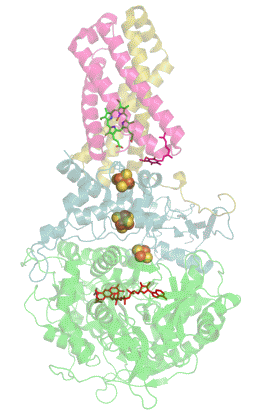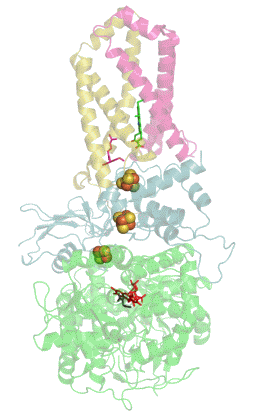
Сукцинатдегидрогеназа или сукцинат-убихинон-оксидорекдуктаза, также известная как комплекс II — белковый комплекс, расположенный во внутренней мембране митохондрий и мембранах многих прокариотических организмов. Одновременно участвует в цикле трикарбоновых кислот и дыхательной цепи переноса электронов.
Общий путь катаболизма — совокупность биохимических процессов, которая включает в себя:
- окисление пирувата до ацетил-КоА;
- окисление ацетил-КоА в цикле трикарбоновых кислот;

Глиоксила́тный цикл, или глиоксила́тный шунт — анаболический путь, имеющийся у растений, бактерий, протистов и грибов, представляет собой видоизменённый цикл трикарбоновых кислот. Глиоксилатный цикл служит для превращения ацетил-СоА в сукцинат, который далее используется для синтеза углеводов. У микроорганизмов он обеспечивает утилизацию простых углеродных соединений в качестве источника углерода, когда более сложные источники, например, глюкоза, недоступны, а также может считаться одной из анаплеротических реакций цикла трикарбоновых кислот, восполняющей количества сукцината и малата. Считается, что у животных глиоксилатный цикл отсутствует, однако в последние годы в некоторых тканях животных были обнаружены ключевые ферменты цикла — малатсинтаза и изоцитратлиаза.

Обра́тный цикл Кре́бса, также известный как обра́тный цикл трикарбо́новых кисло́т, или цикл А́рнона — последовательность химических реакций, которую некоторые бактерии используют для синтеза органических соединений из диоксида углерода и воды.

ГАМК-оперон несет ответственность за превращение γ-аминобутирата (ГАМК) в сукцинат. ГАМК -оперон включает три структурных гена - gabD , gabT и gabP , которые кодируют сукцинат полуальдегида дегидрогеназы, ГАМК-трансаминазы и ГАМК-пермеазы соответственно. Существует регулирующий ген csiR, ниже оперона, который кодирует предполагаемый репрессор транскрипции и активируется при ограничении азота.
ГАМК-шунт — это альтернативный вариант ЦТК, характерный для нервной ткани. Он также функционирует в растениях, где активируется в стрессовых условиях, например при недостатке кислорода.

Фумаратредуктаза — фермент, катализирующий реакцию превращения фумарата в сукцинат. Является важным ферментом для метаболизма микроорганизмов, поскольку участвует в процессе анаэробного дыхания.
- Сукцинат + акцептор ↔ фумарат + восстановленный акцептор
Сукцинил-Кофермент А, сокращенно сукцинил-КоА, представляет собой тиоэфир дикарбоновой янтарной кислоты и кофермента А. Принимает участие во многих биохимических путях, в частности, является интермедиатом цикла Кребса. Служит предшественником синтеза δ-аминолевулиновой кислоты, являющейся первым специфическим интермедиатом в синтезе порфиринов.
Пропионовоки́слое броже́ние — вид брожения, при котором субстрат сбраживается до пропионовой кислоты (пропионата) и уксусной кислоты (ацетата). Пропионовокислое брожение осуществляют преимущественно бактерии подпорядка Propionibacterineae класса Actinobacteria, обитающие в рубце и кишечнике жвачных животных.
ГАМК-трансаминаза (ГАМК-Т) — семейство ферментов трансаминаз, которые разрушают медиатор ГАМК.
