
Рибонуклеи́новая кислота́ (РНК) — одна из трёх основных макромолекул, которые содержатся в клетках всех живых организмов и играют важную роль в кодировании, прочтении, регуляции и экспрессии генов.

Генети́ческий код — совокупность правил, согласно которым в живых клетках последовательность кодонов переводится в последовательность аминокислот (белков). Собственно перевод (трансляцию) осуществляет рибосома, которая соединяет аминокислоты в цепочку согласно инструкции, записанной в кодонах мРНК. Соответствующие аминокислоты доставляются в рибосому молекулами тРНК. Генетический код всех живых организмов Земли един, что свидетельствует о наличии общего предка.

Рибосо́ма — важнейшая немембранная органелла всех живых клеток, служащая для биосинтеза белка из аминокислот по заданной матрице на основе генетической информации, предоставляемой матричной РНК (мРНК). Этот процесс называется трансляцией. Рибосомы имеют сферическую или слегка эллипсоидную форму, диаметром от 15—20 нанометров (прокариоты) до 25—30 нанометров (эукариоты), состоят из большой и малой субъединиц. Малая субъединица считывает информацию с матричной РНК, а большая — присоединяет соответствующую аминокислоту к синтезируемой цепочке белка.

16S рРНК — один из трёх основных типов рРНК, образующих основу рибосом прокариот. Цифры в названии рРНК равны значению константы седиментации. Соответственно, для данной молекулы это значение равно 16S. Всего в прокариотических микроорганизмах обнаружено три типа рРНК: 23S и 5S в большой субъединице рибосомы (50S), 16S в малой субъединице рибосомы (30S). Аналогично, константы двух других молекул рРНК равны 23 и 5 S соответственно. Эукариотическим аналогом 16S рРНК является 18S рРНК.

Нуклеи́новая кислота — высокомолекулярное органическое соединение, биополимер (полинуклеотид), образованный остатками нуклеотидов. Нуклеиновые кислоты ДНК и РНК присутствуют в клетках всех живых организмов и выполняют важнейшие функции по хранению, передаче и реализации наследственной информации.

РНК-полимераза — фермент, осуществляющий синтез молекул РНК. В узком смысле, РНК-полимеразой обычно называют ДНК-зависимые РНК-полимеразы, осуществляющие синтез молекул РНК на матрице ДНК, то есть осуществляющие транскрипцию. Ферменты класса РНК-полимераз очень важны для функционирования клетки, поэтому они имеются во всех организмах и во многих вирусах. Химически РНК-полимеразы являются нуклеотидил-трансферазами, полимеризующими рибонуклеотиды на 3'-конце цепи РНК.

Трансля́ция — осуществляемый рибосомой процесс синтеза белка из аминокислот на матрице информационной (матричной) РНК ; реализация генетической информации.

Ма́тричная рибонуклеи́новая кислота́ — РНК, содержащая информацию о первичной структуре белков. мРНК синтезируется на основе ДНК в ходе транскрипции, после чего, в свою очередь, используется в ходе трансляции как матрица для синтеза белков. Тем самым мРНК играет важную роль в «проявлении» (экспрессии) генов.

Тра́нспортно-ма́тричная РНК, также известна как 10Sa-РНК и SsrA-РНК, — небольшая РНК длиной от 260 до 430 нуклеотидов, которая участвует в высвобождении рибосом, «застрявших» во время трансляции проблемных участков мРНК, а также разрушении получившихся в ходе неполной трансляции дефектных пептидов. Механизм высвобождения рибосомы с дефектной мРНК при участии тмРНК получил название транс-трансляции. Первая тмРНК была обнаружена в 1994 году у кишечной палочки Escherichia coli, и с тех пор тмРНК были описаны у разных групп бактерий. Гены тмРНК обнаруживаются в геномах практически всех бактерий и многих органелл.
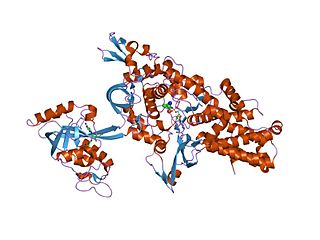
Аминоацил-тРНК-синтетаза (АРСаза) — фермент (синтетаза), катализирующий образование аминоацил-тРНК в реакции этерификации определённой аминокислоты с соответствующей ей молекулой тРНК. Для каждой протеиногенной аминокислоты существует по меньшей мере одна аминоацил-тРНК-синтетаза.
Полиаденили́рование — это процесс присоединения большого количества остатков аденозинмонофосфата к 3'-концу первичной мРНК (пре-мРНК). Иными словами, поли(А)-хвост — это фрагмент молекулы мРНК, азотистые основания которого представлены только аденином. У эукариот полиаденилирование является частью процессинга мРНК — процесса созревания первичного транскрипта в зрелую мРНК, готовую для трансляции. Процессинг, в свою очередь, является одним из этапов экспрессии генов.

Редакти́рование РНК — процесс, в ходе которого нуклеотиды в новосинтезированной РНК подвергаются химическим модификациям. Редактирование РНК также может включать вставку, делецию или замену нуклеотидов в молекуле РНК. Редактирование РНК — довольно редкий процесс, и типичные этапы процессинга мРНК обычно не рассматриваются как редактирование.

Процессинг РНК — совокупность процессов в клетках эукариот, которые приводят к превращению первичного транскрипта в зрелую РНК.

Кэп, 5'-кэп, или кэп-структура — структура на 5'-конце матричных РНК (мРНК) и некоторых других РНК эукариот. Кэп состоит из одного или нескольких модифицированных нуклеотидов и характерен только для транскриптов, синтезируемых РНК-полимеразой II. Наличие кэпа — один из признаков, отличающих эукариотические мРНК от прокариотических, которые несут трифосфат на 5'-конце. Это и другие отличия обуславливают существенно более высокую стабильность, особый механизм инициации трансляции и другие особенности жизненного цикла эукариотической мРНК.

Шпи́лька — в молекулярной биологии элемент вторичной структуры РНК, а также одноцепочечной ДНК. Шпилька образуется в том случае, когда две последовательности одной и той же цепи комплементарны друг другу и соединяются друг с другом, перегибаясь одна к другой и образуя на конце неспаренный участок — петлю. Такие комплементарные последовательности нередко представляют собой палиндромные последовательности.
Предсказа́ние втори́чной структу́ры РНК — метод определения вторичной структуры нуклеиновой кислоты по последовательности её нуклеотидов. Вторичную структуру можно предсказывать для единичной последовательности или анализировать множественное выравнивание семейства родственных РНК.
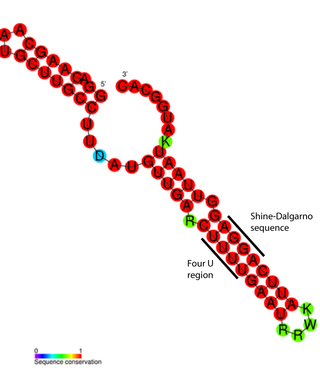
РНК-термо́метр, или РНК-термосе́нсор — температурочувствительная некодирующая РНК, которая принимает участие в регуляции экспрессии генов. РНК-термометры, как правило, регулируют гены, которые необходимы для ответа на тепловой или холодовой шок, однако показано их участие в регуляции длительного голодания и патогенности.
Трансля́ция у прокарио́т — процесс синтеза белка на матрице мРНК, происходящий в клетках прокариотических организмов. В отличие от аналогичного процесса у эукариот, в трансляции у прокариот принимает участие рибосома 70S, а первой (инициаторной) аминокислотой выступает формилметионин, а не метионин.

Nonstop-деграда́ция — механизм контроля качества мРНК, направленный на выявление мРНК, лишённых стоп-кодона, и предотвращение их трансляции. В ходе nonstop-деградации рибосома, которая успела значительно продвинуться в сторону 3'-конца мРНК, диссоциирует, и мРНК направляется в экзосомный комплекс или к РНКазе R для дальнейшего разрушения.
Кольцевы́е РНК — тип молекул РНК, концы которых замкнуты друг на друга с помощью ковалентной связи между концевыми нуклеотидами. Кольцевые РНК могут образовываться из интронов или в результате выпетливания различных участков созревающего транскрипта. Хотя кольцевые РНК обычно относят к числу некодирующих РНК, в настоящее время накапливаются свидетельства того, что они могут кодировать пептиды. Конкретные функции кольцевых РНК до конца не изучены, но, вероятно, они задействованы в регуляции экспрессии генов. Кольцевых РНК особенно много в головном мозге, они свободно циркулируют в плазме крови. Возможно, в будущем кольцевые РНК будут использовать в качестве биомаркеров различных видов рака.

