Термиты
| Термиты | |
|---|---|
 Coptotermes formosanus | |
| Научная классификация | |
Домен: Эукариоты Царство: Животные Подцарство: Эуметазои Без ранга: Двусторонне-симметричные Без ранга: Первичноротые Без ранга: Линяющие Без ранга: Тип: Членистоногие Подтип: Трахейнодышащие Надкласс: Шестиногие Класс: Насекомые Подкласс: Крылатые насекомые Инфракласс: Новокрылые Клада: Надотряд: Тараканообразные Отряд: Таракановые Инфраотряд: Термиты | |
| Международное научное название | |
| Isoptera Brullé, 1832 | |
| Семейства | |
| |

Терми́ты, или белые муравьи[1][2] (лат. Isoptera), — инфраотряд[3][4] общественных насекомых с неполным превращением, родственных тараканам. Несмотря на название «белые муравьи», не являются муравьями, хотя во многом и схожи с ними по образу жизни.
Долгое время термитов считали самостоятельным отрядом (2009)[5], в последнее время их таксономический статус дискутируется и рассматривается в ранге от инфраотряда (2011, 2013)[3][4] до эписемейства (Termitoidae; 2007) в составе тараканообразных[6][7]. Как муравьи и медоносные пчёлы, термиты живут большими семьями с разделением труда между членами колонии и наличием различных каст (солдаты, рабочие, няньки, королева, король и другие). Питаются, главным образом, растительным материалом, мёртвой древесиной, содержащими целлюлозу, для переваривания которой содержат в кишечном тракте множество симбионтных микроорганизмов. В тропиках и субтропиках играют важную роль в почвообразовании. Термиты являются деликатесом в рационе некоторых народов и используются во многих традиционных лекарствах. Численность семей достигает нескольких миллионов особей, а продолжительность жизни некоторых королев превышает несколько десятков лет. Несколько сотен видов экономически значимы в качестве вредителей, которые могут нанести серьёзный ущерб зданиям, сельскохозяйственным культурам или лесопосадкам. Некоторые виды, такие как Cryptotermes brevis, считаются инвазивными видами[8]. В мире известно 2933 современных вида термитов (данные на 2013 год; вместе с ископаемыми их в сумме 3106 видов)[4].
Общие сведения
Как и все общественные насекомые, особи термитов чётко делятся на три основных группы: рабочих особей, особей-солдат и особей, способных к половому размножению. Рабочие термиты имеют мягкое белое тело, как правило, менее 10 мм в длину. Глаза редуцированы или отсутствуют. В противоположность им, репродуктивные особи имеют тёмное тело и развитые глаза, а также две пары длинных треугольных крыльев, которые, однако, сбрасываются после единственного в жизни репродуктивной особи полёта. Обладают свойством эусоциальности[9][10].
Как группа термиты произошли от таракановых в триасовом периоде, на основании чего некоторые энтомологи включают термитов в отряд тараканообразных. Тараканы рода Cryptocercus, знаменитые своей развитой для тараканообразных заботой о потомстве, носят в кишечнике микрофлору, схожую с таковой у термитов, а среди термитов имеется примитивный вид Mastotermes darwiniensis, по характеристикам близкий как к тараканам, так и к другим термитам. Неизвестно, каким именно путём термиты пришли к общественному образу жизни, уникальному среди насекомых с неполным превращением, но известно, что ранние термиты были крылатыми и имели сходный между собой облик. Останки термитов довольно часто встречаются в янтарях разных периодов.
Тела взрослых термитов и их крылья окрашены в различные цвета от беловато-жёлтого до чёрного. Головы солдат могут быть светло-жёлтыми, оранжевыми, красновато-коричневыми или чёрными. Среди самых маленьких по размеру солдат термитов представители вида Atlantitermes snyderi (Nasutitermitinae) из Тринидада и Гайаны (Южная Америка) длиной 2,5 мм, а среди крупнейших — солдаты Zootermopsis laticeps (Termopsidae) из Аризоны (США) и Мексики длиной 22 мм. Крупнейшими среди крылатых половых особей являются самки и самцы африканского рода термитов Macrotermes, чья длина вместе с крыльями достигает 45 мм, а среди мельчайших крылатые термиты Serritermes serrifer (Serritermitidae) — 6 мм с крыльями. Крылатые особи некоторых представителей Incisitermes и Glyptotermes (Kalotermitidae) и Apicotermitinae имеют длину менее 7 мм с крыльями. Численность семей варьируется от нескольких сотен (Kalotermitidae) до нескольких миллионов особей (Rhinotermitidae, Termitidae)[11]. Вес взрослых бескрылых термитов Mastotermes достигает 52 мг[12].
Термиты распространены в тропических и субтропических регионах и насчитывают более 2900 современных видов[13][14]. В России обитают два вида термитов (в районе Сочи и Владивостока). На территории СНГ обитают 7 видов термитов, из них 4 вида, в том числе — туркестанский (Anacanthotermes turkestanicus) и большой закаспийский (Anacanthotermes ahngerianus), — причиняют значительный вред хозяйству[15].
Распространение
Термиты обнаружены на всех континентах, кроме Антарктиды. Меньше всего они представлены в Европе (10 видов двух родов Kalotermes и Reticulitermes) и в Северной Америке (50 видов), в то время как в Южной Америке обитает более 400 видов[16], а в Африке около 1000 видов. В одном только Национальном парке Крюгера (ЮАР) нашли около 1,1 млн активных термитников[17]. В Азии встречается 435 видов термитов, главным образом в Китае, южнее реки Янцзы[16]. В Австралии более 360 видов термитов[16]. Из-за своей тонкой кутикулы термиты бедно представлены в холодном и умеренном климате[16][18]. Один инвазивный американский вид (Cryptotermes brevis) завезён в Австралию[19].
| Азия | Африка | Северная Америка | Южная Америка | Европа | Австралия | |
|---|---|---|---|---|---|---|
| Число видов | 435 | 1,000 | 50 | 400 | 10 | 360 |
Эндемизм
Примеры родового эндемизма. Эндемики Неотропики — Acorhinotermes, Cornitermes, Cylindrotermes, Glossotermes, Dolichorhinotermes, Labiotermes, Paracornitermes, Procornitermes, Rhinotermes, Syntermes и другие (63 эндемичных рода, 78 % от всех родов Неотропики). Эндемики Карибских островов — Antillitermes, Caribitermes, Parvitermes. Gnathamitermes и Zootermopsis встречается в Неарктике (США, Канаде и Мексике), Hoplotermes и Marginitermes — в Мексике. Drepanotermes и Mastotermes— в Австралии; Stolotermes обнаружен в ЮАР и в Австралии и Океании (Новая Зеландия, Тасмания). Эндемики острова Новая Гвинея — Diwaititermes и Niuginitermes. Эндемики Африки и другие (85 эндемичных родов, 77 % от всех родов Афротропики) — Batillitermes, Cephalotermes, Cubitermes, Lepidotermes, Noditermes, Ovambotermes, Procubitermes, Promirotermes, Pseudomicrotermes, Thoracotermes, Tuberculitermes, Unicornitermes. На Шри-Ланке два эндемичных рода Ceylonitermes и Ceylonitermellus. Эндемики Мадагаскара — Capritermes, Quasitermes. Эндемики Малайского полуострова и Калимантан — Orientotermes, Prohamitermes, Protohamitermes. В Индо-Китае и на Суматре встречается Globitremes. Pseudhamitremes известен из Камбоджи. Serritermitidae встречаются только в Бразилии. Termitogeton (Termitogetoninae) обнаружен на Малайском полуострове, Суматре, Калимантан и Шри-Ланке. Stylotermes (Stylotermitininae) обнаружен в Индии, Индо-Китае, Китае и на Калимантан. Psammotermes встречается в пустынях Африки, на Ближнем Востоке и в Индии[20][21].
Структура колонии и поведение
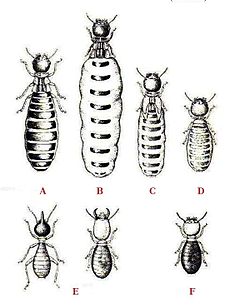
A — Правящий король
B — Правящая царица
C — Вторая царица
D — Третья царица
E — Солдаты
F — Рабочий
Как и все общественные насекомые, термиты живут в колониях, число зрелых особей в которых может достигать от нескольких сотен до нескольких миллионов и состоящих из каст. Типичная колония состоит из личинок (нимф), рабочих, солдат и репродуктивных особей. Сооружение термитов — термитник. В отличие от муравьёв, а также ос и пчёл, у наиболее эволюционно продвинутых видов термитов кастовая принадлежность задаётся генетически. У более примитивных видов кастовая принадлежность особи зависит от того, чем её кормят другие термиты в период развития и какие феромоны они при этом выделяют[22].
Бесполое размножение в виде телитокического партеногенеза обнаружено у 7 видов термитов, в том числе: Reticulitermes speratus, Zootermopsis angusticollis, Zootermopsis nevadensis, Kalotermes flavicollis, Bifiditermes beesoni. Популяции вида Glyptotermes nakajimai с островов Кюсю и Сикоку размножается исключительно партеногенетически[23]. Диплоидный хромосомный набор термитов составляет 2n=28—56[24], а у примитивного Mastotermes darwiniensis 2n=96[25][26].
Репродуктивные особи
Среди репродуктивных особей в гнезде выделяют короля и королеву. Это особи, уже утратившие крылья и иногда глаза и выполняющие репродуктивную функцию в гнезде. Достигнувшая зрелости королева может откладывать несколько тысяч яиц в день, превращаясь в своеобразную «фабрику яиц». В этом состоянии её грудь и особенно брюшко увеличиваются, делая королеву в несколько десятков раз крупнее любого рабочего термита (10 см и больше). Из-за гигантского брюшка королева утрачивает способность к самостоятельному передвижению, поэтому когда возникает необходимость переместить её в другую камеру колонии, сотни рабочих объединяются, чтобы перенести её. На поверхности тела королевы выделяются особые феромоны, слизываемые рабочими, которые способствуют объединению колонии. У некоторых видов эти феромоны оказываются настолько притягательными для рабочих, что они вгрызаются жвалами в брюшко королевы (однако к её гибели это приводит крайне редко).
В камере королевы находится король, который лишь немного больше рабочего термита. Он продолжает спариваться с самкой на протяжении жизни, в отличие от, например, муравьёв, у которых мужские особи гибнут сразу после спаривания, а сперма хранится внутри королевы (матки) в придатках яичников.

Остальные репродуктивные особи обладают крыльями и служат для создания новых колоний. В определённое время года они вылетают из гнезда и спариваются в воздухе, после чего самец и самка, спустившись на землю, отгрызают себе крылья и вдвоём основывают новую колонию. У некоторых видов термитов неполовозрелые репродуктивные особи составляют подкасту, предназначенную для замены короля и королевы в случае их гибели. Однако происходит это крайне редко.
У спаривающихся пар термитов наблюдается поведение тандемного бега. После вылета крылатых взрослых особей из своих гнезд, самки и самцы садятся на землю, сбрасывают крылья и бегут в поисках партнера для спаривания. При встрече пара совершает тандемный бег в поисках подходящего места для основания колонии[27].
Учёные Университета Ямагути и Университета Тоттори пришли к выводу, что королевы термитов Reticulitermes speratus живут дольше, чем рабочие, за счёт повышенной активности генов, отвечающих за кодирование ферментов-антиоксидантов: каталазы и семьи пероксиредоксинов[28].
Рабочие
В отличие от муравьёв, среди рабочих и солдат термитов поровну самок и самцов. Рабочие термиты занимаются фуражированием, хранением пищи, заботой о потомстве, строительством и ремонтом колонии. Рабочие — единственная каста, способная переваривать целлюлозу благодаря особым кишечным микроорганизмам-симбионтам. Именно они занимаются кормлением всех остальных термитов. Своими впечатляющими характеристиками колонии также обязаны рабочим.


Стены колонии строятся из комбинации экскрементов, измельчённой древесины и слюны. В гнезде предусмотрены места для разведения грибковых садов, содержания яиц и молодых личинок, репродуктивных особей, а также обширная сеть вентиляционных туннелей, которые позволяют поддерживать внутри термитника практически постоянный микроклимат. Кроме того, иногда имеются также помещения для термитофилов — животных, сосуществующих с термитами в симбиозе.
Солдаты

Солдаты — особая каста рабочих особей, которая имеет анатомические и поведенческие специализации, прежде всего против нападения муравьёв. Многие имеют челюсти столь увеличенные, что неспособны самостоятельно питаться. Солдаты тропических видов термитов-носорогов имеют особый вырост на голове, через который выстреливают защитной жидкостью. У термитов, прогрызающих ходы в дереве, солдаты обычно имеют широкие головы, которые позволяют блокировать узкие туннели и не допустить дальнейшего проникновения врага в гнездо. Когда нарушена целостность стен термитника и ситуация такова, что требует вмешательства больше чем одного солдата, солдаты образуют защитное формирование, напоминающее фалангу, и начинают беспорядочно нападать на свою жертву, в то время как рабочие заделывают отверстие. Как правило, солдаты фаланги впоследствии сами становятся жертвой муравьев, поскольку после восстановления стены термитника фаланга лишается возможности вернуться в термитник.
Химическая защита отсутствует в семействах Termopsidae, Hodotermidae и Kalotermitidae. Слюнные или лабиальные железы развиты у всех термитов и всех каст. У части видов солдаты продуцируют защитные секреты, например, у Mastotermes и многих Macrotermitinae. Гипертрофированная фронтальная слюнная железа представлена и у части Termitinae с щёлкающими мандибулами: по крайней мере у Dentispicotermes секреты железы выводятся путём разрыва железы (autothisis, или саморазрыв стенок тела). Эта уникальная в мире насекомых непарная железа является синапоморфией в семействах Rhinotermitidae, Serritermitidae и Termitidae. Малозаметная у рабочих и очень маленькая у половых имаго, эта железа максимально развита у солдат и продуцирует разнообразные химические компоненты. Их выброс производится через специализированное отверстие на голове, которое называется фонтанелла (фронтальная пора). У некоторых термитов фронтальная пора закрыта и поэтому секреты железы выводятся путём разрыва всей железы и брюшка (аутотизис), например у Serritermes и Globitermes[20]. Термиты Globitermes sulphureus, известные как «термиты-камикадзе», используют форму суицидального альтруизма, известного как аутотизис, в качестве защитного механизма[29].
Защитная фронтальная железа выбрызгивает защитные секреты во врага через выводные протоки в голове касты солдат (у носатых термитов этот выход находится в специальной фронтальной поре в специализированном «носу» головной капсулы). Фронтальная железа сильно развита в брюшке у солдат Rhinotermitidae (Coptotermes, Psammotermes, Reticulitermes, Prorhinotermes, Schedorhinotermes) и в голове солдат Termitidae (развиты у Nasutitermitinae, но редуцированы в подсемействах Macrotermitinae и Termitinae, которые защищаются мощными мандибулами)[30].
Число солдат в колонии зависит от активности семьи и обычно составляет несколько процентов от всего населения. У меньшинства видов доля солдат менее 3 %. Около 4-6 % — у видов родов Cryptotermes (Nutting, 1970; Bouillon, 1970), Incisitermes (Harvey, 1934; Nutting, 1970), Kalotermes (Harris, 1954; Grasse and Noirot, 1958) и Glyptotermes (Danthanarayana and Fernando, 1970). Около 1-9 % у видов родов Neolermes (Nagin, 1972; Sen-Sarma and Mishra, 1972), Stolotermes (Morgan, 1959), Odontotermes (Josens, 1974) и Macrotermes (Pangga, 1936). По данным Josens (1972, 1974), собранным на Берегу Слоновой Кости (Западная Африка), пропорция солдат варьирует в пределах 12-16 % у видов Basidentermes potens, Promirotermes holmgreni, Ancistrotermes cavithorax, Microtermes toumodiensis, Pseudacanthotermes militaris. В колониях носатых термитов (в двух подсемействах: Coptotermitinae и Nasutitermitinae) у которых фронтальная железа наиболее развита, пропорция солдат может быть ещё выше. У видов Coptotermes formosanus и Coptotermes vastator, солдат около 10 % (Smythe and Mauldin, 1972; King and Spink, 1974, 1975; Pangga, 1936), в то время как у Hospitalitermes hospitalis (Pangga, 1936) и различных видов рода Nasutitermes (Kfecek, 1970; Gay and Wetherly, 1970; Holdaway et al., 1935) число носатых солдат составляет около 10 % от всей активной части популяции. Они составляют 15 —21 % у вида Trinervitermes geminatus (Bouillon, 1970; Josens, 1974), 20 % у видов Trinervitermes togoensis (Josens, 1974) и Nasutitermes spp. (Pangga, 1936; Gay et al., 1955), 25 % у термитов Tenuirostritermes tenuirostris (Weesner, 1953), и даже до 30 % у термитов в лабораторных гнёздах Nasutitermes costalis (Hrdy and Zeleny, 1967). Это может быть связано с мелкими размерами солдат носатых термитов, сравнимых с размерами рабочих[30]. У термита — ксилофага Cephalotermes (Termitinae) численность солдат составляет лишь 0,2 %[20]. Потеря касты солдат наблюдается в двух подсемействах семейства термитов Termitidae. У Apicotermitinae более чем половина африканских видов не имеют солдат, так же как и все неотропические роды. Наоборот, все юговосточноазиатские роды имеют солдат, хотя эта каста у них очень редка (род Speculitermes долгое время считался не имеющим солдат)[20]. В подсемействе Termitinae только в 3 родах нет касты солдат: Protohamitermes и Orientotermes (два близких таксона из Ориентальной области)[31] и Invasitermes[32] из Австралии[20].
Биология и экология
Локомоция и фуражировка
Поскольку касты рабочих и солдат не имеют крыльев и, следовательно, никогда не летают, а репродуктивные особи используют крылья только в течение короткого промежутка времени, термиты в основном полагаются на ноги для передвижения[33].
Поведение при добывании пищи зависит от вида термитов. Например, некоторые из них питаются древесными структурами, в которых они обитают, а другие собирают пищу рядом с гнездом[34]. Большинство рабочих редко выбираются на открытый воздух и не добывают корм без защиты; они полагаются на защитное покрытие и пути отступления, чтобы защитить себя от хищников[35]. Подземные термиты строят туннели в поисках пищи, а рабочие, которым удаётся найти источники пищи, вербуют соплеменников по гнезду, выпуская феромоны, привлекающие рабочих[36]. Рабочие-фуражиры используют семиохимические вещества для общения друг с другом[37], а рабочие, которые начинают добывать корм за пределами своего гнезда, выделяют феромоны из своих стернальных желёз[38]. Например, у вида Nasutitermes costalis экспедиция за кормом состоит из трёх этапов: во-первых, солдаты исследуют территорию. Когда они находят источник пищи, они привлекают других солдат и небольшую группу рабочих. На втором этапе рабочие приходят в большом количестве на это место. Третий этап отмечен уменьшением количества присутствующих солдат и увеличением числа рабочих[39]. Изолированные рабочие-термиты могут использовать случайное блуждание как оптимизированную стратегию для поиска своих соплеменников или поиска пищи[40].
По наличию или отсутствию массовой фуражировки термитов классифицируют на внутригнездовые и фуражирующие (с массовыми фуражировочными колоннами и множественными гнездовьями). К внутригнездовым относятся Zootermopsis, все роды Stolotermitidae, Stylotermitidae и Serritermitidae, почти все роды Kalotermitidae и ринотермитиды Prorhinotermes и Termitogeton. К фуражирующим относятся Mastotermes, Hodotermopsis, все Hodotermitidae, Paraneotermes, большинство родов Rhinotermitidae и все роды Termitidae[41].
Конкуренция
Конкуренция между двумя колониями всегда приводит к агонистическому поведению в отношениях друг с другом, что выражается в массовых боях. Эти бои могут вызывать гибель обеих сторон и, в некоторых случаях, прирост или потерю территории[42][43]. У отдельных видов даже образуются кладбища в виде «могильных ямок» («Cemetery pits»), где складированы (похоронены) тела мёртвых термитов[44].
Исследования показывают, что когда термиты сталкиваются друг с другом в районах фуражировки, некоторые из них намеренно блокируют проходы, чтобы предотвратить проникновение других термитов[45][46]. Мёртвые термиты из других колоний, обнаруженные в поисковых туннелях, приводят к изоляции этого участка и, следовательно, к необходимости создания новых туннелей[47]. Конфликт между двумя конкурентами происходит не всегда. Например, хотя они и могут блокировать друг друга, колонии Macrotermes bellicosus и Macrotermes subhyalinus не всегда проявляют агрессию по отношению друг к другу[48]. Суицидное поведение обнаружено у вида Coptotermes formosanus. Иногда две разные колонии C. formosanus могут обнаружить один и тот же кормовой ресурс и вступить в физический конфликт. При этом некоторые термиты плотно втискиваются в фуражировочные ходы и там умирают, успешно блокируя туннель и заканчивая все агонистические взаимодействия двух колоний[49].
Среди представителей репродуктивной касты неотенические самки (будущие матки) могут конкурировать друг с другом, чтобы стать доминирующей королевой, когда нет первичных яйцекладущих особей (королевы, или самки-основательницы колонии). Эта борьба между молодыми королевами приводит к гибели их всех, кроме единственной королевы, которая с основным самцом (королём) берёт на себя главную яйцекладущую функцию колонию[50].
Муравьи и термиты могут конкурировать друг с другом за место гнездования. В частности, муравьи, которые охотятся на термитов, обычно отрицательно влияют на гнездящиеся на деревьях виды Isoptera[51].
Коммуникация
Большинство термитов слепы, поэтому их общение происходит главным образом с помощью химических, механических и феромонных сигналов[45][52]. Эти способы коммуникации используются во множестве видов деятельности, в том числе во время фуражировки, обнаружении половых каст, строительства гнёзд, распознавании соплеменников, во время брачного полёта, обнаружения и борьбы с врагами и при защите гнёзд[45][52]. Самый распространённый способ общения термитов — через контакт антенн (усиками)[45]. Известно несколько феромонов, в том числе контактные феромоны (которые передаются, когда рабочие занимаются трофаллаксисом или грумингом[англ.]) и феромоны тревоги, следовые феромоны и половые. Феромоны тревоги и другие защитные химикаты секретируются из фронтальной железы. Следовые феромоны выделяются из стернальной железы, а половые феромоны продуцируются из двух железистых источников: стернальной и тергальной желёз[52]. Когда термиты выходят на поиски пищи, они фуражируют колоннами по поверхности почвы через растительные заросли. Тропинка может быть идентифицирована фекальными отложениями или углублёнными дорожками. Рабочие оставляют на этих тропах феромоны, которые обнаруживаются другими соплеменниками с помощью обонятельных рецепторов[53]. Термиты могут также взаимодействовать через механические сигналы, вибрации и физический контакт[45][53]. Эти сигналы часто используются для связи особей при тревоге (тревожная коммуникация) или для оценки источника питания[45][54].
Когда термиты строят свои гнёзда, они используют преимущественно косвенную связь. Ни один термит не отвечает за какой-либо конкретный фрагмент строительства. Отдельные термиты реагируют на конкретную ситуацию, но на групповом уровне они демонстрируют своеобразное «коллективное сознание». Конкретные структуры или другие объекты, такие как гранулы почвы или столбов, заставляют термитов начинать процесс постройки. Термит добавляет эти объекты в существующие структуры, и такое поведение способствует строительному поведению у других рабочих. Результатом является самоорганизованный процесс, при котором информация, которая направляет деятельность термитов, является результатом изменений в окружающей среде, а не прямого контакта между индивидуумами[45].
Термиты могут отличать соплеменников от чужаков через химическую коммуникацию и кишечных симбионтов: химические вещества, состоящие из углеводородов, высвобождаемых из кутикулы, позволяют распознавать чужеродных термитов[55][56]. Каждая колония имеет свой особый запах. Этот запах является результатом генетических и экологических факторов, таких как диета термитов и состав бактерий в кишечнике термитов[57].
Защита от врагов
Для защиты колонии термиты полагаются на сигналы тревожной связи[45]. Феромоны тревоги могут выделяться, когда гнездо нарушено или подвергается нападению врагов или потенциальных патогенов. Термиты всегда избегают соплеменников, заражённых спорами энтомопатогенных грибов Metarhizium anisopliae, благодаря вибрационным сигналам, издаваемым заражёнными термитами[58]. Другие методы защиты включают интенсивные подёргивания телом, выделение липкой раздражающей жидкости из лобной железы и испражнения, содержащие феромоны тревоги[45][59].
Взаимоотношения с другими организмами
Многочисленные виды организмов взаимосвязаны с термитами в различных формах отношений от паразитизма и хищничества до облигатного симбиоза и термитофилии. Известен вид грибка, который имитирует яйца термитов, успешно избегая своих естественных хищников. Эти маленькие коричневые шарики, известные как «термитные шарики», которые сами термиты редко убивают, и в некоторых случаях рабочие ухаживают за ними[60]. Этот грибок не только имитирует яйца, но и вырабатывая особый фермент для переваривания целлюлозы, известный как глюкозидаза[61]. Уникальное имитационное поведение существует между различными видами жуков-стафилинид рода Trichopsenius (Trichopseniini) и некоторыми видами термитов Reticulitermes. Жуки имеют те же кутикулярные углеводороды, что и термиты, и даже биосинтезируют их. Эта химическая мимикрия позволяет жукам интегрироваться в колонии термитов[62]. Развитые придатки на физогастрически увеличенном брюшке Austrospirachtha mimetes (Aleocharinae) позволяют этому жуку имитировать форму рабочего термита[63].
Эволюция термитофилии у жуков-стафилинид связана со стратегиями гнездования термитов. Для сравнения гнездящихся в одном гнездовом месте видов, вся жизнь колонии которых завершается в одном куске дерева, с фуражирующими видами, которые используют несколько физически разделенных источников пищи и массовые фуражировочные колонны, была построена эпидемиологическая модель. Она предсказывает, что характеристики, связанные с фуражированием (например, увеличенная продолжительность жизни колонии и частые взаимодействия с другими колониями), увеличивают вероятность паразитизма термитофилов. Обнаружено, что фуражирующие виды с большей вероятностью содержат термитофильных стафилинид, чем цельногнездовые виды: 99,6 % известных термитофильных видов были связаны с фуражирующими термитами, тогда как 0,4 % были связаны с цельногнездовыми виды. Филогенетический анализ подтвердил, что термитофилия в первую очередь развилась с фуражирующими термитами[41].
Гнёзда
Гнёзда термитов называются термитниками и, как правило, выглядят в виде крупных холмиков, возвышающихся над поверхностью земли[64]. Основной его функцией является защита термитов от врагов, сухости и жары[65].
Холмики термитников, расположенные в районах с ливневыми и непрерывными осадками, подвергаются риску эрозии своей структуры из-за их глиняной основной конструкции. Те гнёзда, которые сделаны из картона (пережёванной растительной и в основном древесной массы), могут обеспечить защиту от дождя и фактически выдерживают сильные осадки[64]. Определённые области в термитниках используются в качестве силовых точек в случае нарушения или пролома гнезда. Например, колонии Cubitermes строят узкие туннели, используемые в качестве таких силовых точек, так как диаметр туннелей достаточно мал, чтобы их блокировали солдаты[66]. Высокозащищенная камера, известная как «королевская камера», содержит королеву и короля и используется как последняя линия обороны[65].
Виды рода Macrotermes, возможно, строят самые сложные структуры в мире насекомых, сооружая огромные насыпи[64]. Эти термитники являются одними из крупнейших в мире, достигающими высоты от 8 до 9 м, и состоят из многочисленных ходов, вершин и хребтов[53]. Другой вид термитов, Amitermes meridionalis, может строить гнёзда высотой от 3 до 4 м и шириной 2,5 м. Самый высокий термитник, который когда-либо регистрировался, составлял в высоту 12,8 м и обнаружен в Экваториальной Африке в Демократической Республике Конго[67].
Некоторые термиты строят холмики со сложной видоспецифичной структурой. Например, термиты рода Amitermes (Amitermes meridionalis и A. laurensis) строят «компасные» или «магнитные» холмики, ориентированные с севера на юг[68][69]. С помощью экспериментов было показано, что эта компасная ориентация помогает терморегуляции. Ориентация с севера на юг приводит к тому, что внутренняя температура насыпи быстро увеличивается в течение утра, избегая перегрева от полуденного солнца. Затем температура остаётся на необходимом термитам высоком уровне (на графическом плато) весь оставшийся день до вечера[70].
 Термитник (до 5 м), Австралия
Термитник (до 5 м), Австралия «Компасные» или «магнитные» термитники (Amitermes) ориентированы с севера на юг для избегания полуденной жары
«Компасные» или «магнитные» термитники (Amitermes) ориентированы с севера на юг для избегания полуденной жары Термитник, Австралия
Термитник, Австралия Термиты в гнезде, Мадагаскар
Термиты в гнезде, Мадагаскар Термитник, Намибия
Термитник, Намибия
Хищники

Термиты служат добычей большому разнообразию хищников. Только один вид термитов, например, Hodotermes mossambicus, обнаружен в желудках 65 видов птиц и 19 видов млекопитающих[71], ими питаются муравьи[72][73], членистоногие, рептилии, земноводные, пчёлы, многоножки, тараканы, кузнечики, стрекозы, лягушки[74], змеи[75], скорпионы, пауки[76]. Два вида пауков из семейства Ammoxenidae узко специализированы на термитофагии, то есть на поедании термитов[77][78][79]. Среди крупных хищников такие виды, как трубкозуб, земляной волк, муравьедовые, летучие мыши, медведи, кроличьи бандикуты, многие птицы, ехидны, лисы, галаговые, сумчатый муравьед, мыши и панголины[77][80][81][82]. Земляной волк является насекомоядным млекопитающим, который в основном питается термитами. Он находит свою пищу по звуку, а также обнаруживает запах, выделяемый солдатами. Одна особь земляного волка способна потреблять тысячи термитов за одну ночь, используя свой длинный липкий язык[83][84]. Медведи-губачи разрушают гнёзда термитов, чтобы потреблять их обитателей, в то время как шимпанзе используют палочки как орудия труда, чтобы удить ими насекомых из их гнезда. Анализ структуры костных инструментов, используемых ранним гоминидом Paranthropus robustus, предполагает, что они использовали эти инструменты для выкапывания термитов[85].

Среди всех хищников муравьи рассматриваются главными врагами термитов[72][73]. Некоторые муравьиные роды — специализированные термитофаги. Например, Megaponera охотятся, как правило, исключительно на термитов, проявляя при этом рейдовую активность по захвату добычи, длящуюся по несколько часов[86][87]. Paltothyreus tarsatus представляют собой другой пример рейдера-захватчика термитов, когда каждый муравей уносит в своих челюстях столько особей термитов, сколько может схватить их мандибулами, кроме того, производится мобилизация дополнительных сил муравьёв с помощью химических следовых феромонов[72]. Малайзийский муравей Eurhopalothrix heliscata применяет иную стратегию во время охоты на термитов: прячась в узких закоулках их древесных термитников, он схватывает жертву мгновенно защёлкивающимися жвалами[72]. Tetramorium uelense специализирован в охоте на мелкие виды термитов. Группы отмобилизованных рабочих муравьёв примерно из 10-30 рабочих прибывают в область нахождения термитов, убивая их с помощью своего жала[88]. Муравьи родов Centromyrmex и Iridomyrmex иногда устраивают свои гнёзда в термитниках и в них же охотятся на термитов. Никаких доказательств в пользу иных (кроме хищничества) взаимоотношений не установлено[89][90]. Другие муравьи, включая представителей родов Acanthostichus, Camponotus, Crematogaster, Cylindromyrmex, Leptogenys, Odontomachus, Ophthalmopone, Pachycondyla, Rhytidoponera, огненные муравьи и Wasmannia, также охотятся на термитов[72][80][91]. В отличие от всех этих видов муравьёв, кочевники рода Dorylus редко потребляют термитов, несмотря на всё огромное разнообразие добычи муравьёв-кочевников[92].
Муравьи не единственные беспозвоночные, которые совершают рейды. Многие роющие осы и несколько видов других ос, включая Polybia и Angiopolybia pallens, как обнаружено, совершают рейдовые полёты на термитники, когда из них вылетают брачные рои самок и самцов термитов[93].
Паразиты
Термиты менее подвержены атакам паразитов, чем пчёлы, осы и муравьи, так как они обычно хорошо защищены в их гнёздах термитниках[94][95]. Тем не менее, термиты заражаются различными паразитами. Среди них паразитические мухи[96], клещи Pyemotes и множество видов круглых червей. Среди нематод представители отряда Rhabditida;[97], а также рода Mermis, Diplogaster aerivora и Harteria gallinarum[98]. Под непосредственной угрозой нападения паразитов колония может мигрировать в новое место[99]. Грибные патогены, такие как Aspergillus nomius и Metarhizium anisopliae представляют серьёзную угрозу для колонии термитов, поскольку они не являются специфичными для хозяина и могут заражать большие части колонии[100][101]; заражение обычно происходит при прямом физическом контакте[102]. Вид M. anispliae как известно, ослабляет иммунную систему термитов. Инфицирование видом A. nomius происходит только при нахождении колонии в стрессовом состоянии[101]. Инквилинизм между разными видами термитов не отмечен[103].
Питание
Все термиты едят целлюлозу в той или иной форме, но именно питающиеся деревом особенно ответственны за нанесение ущерба зданиям. Целлюлоза — богатый источник энергии, но трудноперевариваемый. Среди симбиотических организмов в своём кишечнике термиты полагаются в первую очередь на метамонад рода Trichonympha, которые потребляют оставшиеся после переваривания вещества. Кишечные микроорганизмы, в свою очередь, полагаются на особых бактерий, живущих на их оболочках и производящих некоторые пищеварительные ферменты. Эти отношения — один из самых прекрасных примеров мутуализма среди животных. Большинство «высших» термитов, особенно семейства Termitidae, могут производить и свои целлюлозоперерабатывающие ферменты, однако и они сохраняют в своём кишечнике богатую микрофлору.
Некоторые виды термитов занимаются разведением грибных садов, выращивая в них специализированный гриб Termitomyces, который входит в их рацион питания, на собственных экскрементах. После поедания споры гриба проходят неповреждёнными через кишечник термитов и прорастают в свежих фекалиях.
Взаимодействие с человеком
Примерно 10 % видов термитов являются вредителями, принося потери хозяйству человека, исчисляемые миллиардами долларов в год[104] (до 20 млрд $ в мире[105]). Из-за своих пищевых пристрастий термиты в некоторых регионах стали настоящим бичом деревянных зданий. Их скрытность и тактика поедания древесины, при которой её поверхность выглядит абсолютно неповреждённой, являются причиной того, что их обнаруживают слишком поздно. Опасение вызывают также случайные попадания термитов в квартиры, благодаря чему их ареал продвигается и в те регионы, где они не могут жить открыто из-за климата. Попав в дом, термиты не ограничиваются древесиной: все, в чём высока доля целлюлозы, служит для них потенциальной пищей, в том числе и бумага. Последствия этого порой катастрофические: например, в городах Южной Америки из-за постоянного присутствия термитов редко можно найти книгу старше пятидесяти лет[106].
Термиты стараются не контактировать с «открытым воздухом», так как их кутикула очень тонка и не удерживает влагу. Если им необходимо пересечь открытое пространство, они строят туннели-убежища из спрессованной земли и экскрементов.
Основные предосторожности от проникновения термитов:
- Устранение контакта древесины с землёй, использование бетонного фундамента или стальной основы. Но даже в этом случае термиты посредством туннелей способны достичь дерева; кроме того, известны случаи, когда для этой цели они использовали водопроводные трубы.
- Обработка древесины
- Использование древесины, стойкой к определённым видам термитов
Если термиты уже проникли в здание, проводится их удаление инсектицидами. Другой обычный метод — распыление трёхокиси мышьяка, яда медленного действия, используемого в Австралии с 1930-х годов. Яд будет распределён между особями колонии прежде, чем будут видны изменения, что благоприятствует уничтожению всей колонии целиком.
26 видов термитов (главным образом, из родов Cryptotermes, Heterotermes и Coptotermes) стали инвазивными, распространившись с помощью человеческой коммерции далеко за пределы своего нативного ареала[107].
Термиты как пища


43 вида термитов используются в качестве пищи людьми или идут на корм домашнему скоту[108]. Эти насекомые особенно важны в менее развитых странах, где распространено недоедание, поскольку белок, получаемый от термитов, может помочь улучшить рацион людей. Термитов употребляют в пищу во многих регионах мира, но только в последние годы это стало популярным в развитых странах[108].
Термиты потребляются людьми в пищу во многих и разных культурах по всему миру. В Африке крылатые термиты являются важным фактором в диетах местных народов[109]. Разные племена используют разные способы сбора или культивирования насекомых. Иногда люди этих племён собирают солдат термитов нескольких видов. Королевы термитов, хотя их труднее найти, считаются деликатесом[110]. Крылатые термиты имеют высокий уровень привлекательности, так как содержат достаточный запас жира и белка. Они считаются приятными по вкусу, с ореховым привкусом после их приготовления[109].
Крылатые термиты собираются тогда, когда начинается сезон дождей. Во время брачного полёта они обычно видны вокруг электроламп, к свету которых они привлекаются, и поэтому сетки обычно устанавливаются на лампах, а собранные крылатые термиты позже собираются из них. Крылья удаляются с помощью техники, похожей на веялки. Наилучший результат достигается, когда термиты слегка обжареные на горячей плите или жареные до хрустящей корочки. Масло не требуется, так как их тела обычно содержат достаточное количество своего масла. Термитов, как правило, едят, когда поголовье скота и племенные культуры ещё не развились или не произвели достаточной пищи, или если запасы продовольствия из предыдущего вегетационного периода ограничены[109].
Кроме Африки, термиты потребляются местными племенами в Азии и Северной и Южной Америке. В Австралии коренные австралийцы знают, что термиты съедобны, но не потребляют их даже в периоды дефицита. Есть несколько объяснений относительно того, почему они так поступают[109][110]. Гнездовые насыпи, сооружаемые термитами являются основными источниками геофагии во многих странах, включая Кению, Танзанию, Замбию, Зимбабве и Южную Африку[111][112][113][114]. Исследователи предполагают, что термиты являются подходящими кандидатами для использования человеком и космического сельского хозяйства, поскольку они содержат большое количество белка и могут быть использованы для превращения несъедобных отходов в потребительские продукты для людей[115].
В культуре
Термиты были известны человеку с глубокой древности. В древнеиндийской самхите Ригведа (около 1350 год до нашей эры) вероятно, является первым литературным источником, в котором термиты упоминаются в качестве разрушителей древесины. В индийском эпосе «Махабхарата» упоминается, что отшельник Чиавана был неподвижен так долго, что превратился в колонну и термиты полностью облепили его глиной. Имеются народные легенды в Того о демоне в термитниках. Греческий географ Павсаний (около 200 год н. э.) упоминает термитов, как «белых муравьёв» на острове Пефнос у берегов Греции. Путешественник Элиан (около 300 год н. э.) описал конусовидные строения индийских термитов. Многократно упоминавшиеся Цицероном, Виргилием, Плинием, Катоном, Варроном и другими римскими философами «solifugae» — мелкие белые, боящиеся света насекомые, вероятно, были термитами. В XV—XVI веках многие европейские путешественники описывали в своих дневниках и отчётах постройки термитов В Африке, Индии, Гвинеи[116].
Бельгийский писатель М. Метерлинк, увлекавшийся энтомологией, в 1926 году опубликовал натурфилософскую книгу «Жизнь термитов» (фр. La Vie des Termites)[117] в которой писал: «Эта древнейшая из известных цивилизация — самая интересная, самая сложная, самая разумная и, в определённом смысле, самая логичная и лучше всего приспособленная к трудностям существования из тех, что появились на Земле до нашей. С некоторых точек зрения, несмотря на свою жестокость, мрачность и часто омерзительность, она стоит выше цивилизации пчёл, муравьёв и самого человека».
Роль в экологии
Термиты вместе с дождевыми червями и муравьями играют важнейшую роль в циркуляции почвенного вещества, кроме того, крылатые особи служат пищей многочисленным хищникам. Полагают также, что освобождаемый термитами в результате их деятельности метан вносит заметный вклад в общее действие парниковых газов. По своей общей биомассе (от 1 г/м3 до более чем 10 г/м3) термиты сравнимы с общей биомассой наземных позвоночных животных[11].
Термиты вместе с муравьями могут увеличивать урожайность в регионах с сухим и жарким климатом (в экспериментальных условиях в Австралии они увеличивают урожай пшеницы на 36 %), где отсутствуют дождевые черви[118].
Охранный статус
По состоянию на 2020 год 2 вида термитов занесены в Красный список угрожаемых видов МСОП (The IUCN Red List of Threatened Species) как виды, находящиеся на грани исчезновения (CR)[119]:
 Glyptotermes scotti — вид термитов из семейства Kalotermitidae, эндемик небольшого острова Силуэт в Сейшельском архипелаге, где известен только по одной точке обнаружения в северной части острова[120].
Glyptotermes scotti — вид термитов из семейства Kalotermitidae, эндемик небольшого острова Силуэт в Сейшельском архипелаге, где известен только по одной точке обнаружения в северной части острова[120].- Procryptotermes fryeri — вид термитов из семейства Kalotermitidae, эндемик атолла Альдабра (государство Сейшельские Острова)[121].
Ещё два вида включены в категорию «Виды, вызывающие наименьшие опасения» (Least concern): Neotermes laticollis, Nasutitermes maheensis[122].
Систематика
В 2024 году признано 13 современных семейств термитов (не считая 7 вымерших)[123]. Ранее традиционно выделялось 7 или 9 семейств термитов. Затем некоторые подсемейства получили статус семейств[124], добавились Stolotermitidae, Stylotermitidae и Archeorhinotermitidae (Engel & Krishna, 2004). В 2009 году было выделено ещё два семейства: Cratomastotermitidae и Archotermopsidae (Engel, Grimaldi & Krishna, 2009)[5]. В 2021 году было признано 17 семейств термитов (включая ископаемые группы) и проведена реклассификация низших термитов и выделено несколько новых таксонов: †Arceotermitidae, †Krishnatermitidae, †Melqartitermitidae, †Mylacrotermitidae, †Tanytermitidae и другие[125]. В 2024 году дополнительно выделены семейства Heterotermitidae, Psammotermitidae и Termitogetonidae (признано 13 современных семейств термитов и 8 вымерших), а также подсемейства Crepititermitinae, Cylindrotermitinae, Forficulitermitinae, Neocapritermitinae, Protohamitermitinae и Promirotermitinae[123]. Наибольшее число эндемичных родов встречается в следующих регионах: Неотропика (63, 78 %), Афротропика (85, 77 %), Австралазия (23, 64 %), Ориентальная область (56, 61 %)[20]. В мире известно 3106 видов и 331 род (в том числе ископаемых: 173 и 51 (2013)[4].
Палеонтология
Древнейшие ископаемые остатки термитов известны из раннего мела Забайкалья[5] и среднемелового бирманского янтаря[126].
- † Pabuonqedidae
- †Anisotermes xiai Zhao et al., 2019, мел (альб-сеноман), бирманский янтарь
- †Archeorhinotermes rossi Krishna & Grimaldi, 2003, мел (сеноман), бирманский янтарь
- †Baissatermes lapideus Engel, Grimaldi and Krishna, 2007, мел (Байсса, Зазинская свита)
- †Idanotermes desioculus Engel, 2008, эоцен, балтийский янтарь
- †Kachinitermopsis burmensis Poinar, 2009, мел (альб-сеноман), бирманский янтарь
- †Mastotermes monostichus
- †Meiatermes cretacicus Bezerra et al., 2022
- †Morazatermes krishnai Engel & Delclòs, 2010, мел (апт), испанский янтарь
- †Nanotermes isaacae, эоцен
- †Pabuonqed eulna V RŠANSKÝ , 2019, мел, бирманский янтарь
- †Parastylotermes krishnai, эоцен
- †Santonitermes chloeae Engel, Nel & Perrichot, 2011, мел (альб-сеноман), французский янтарь
- †Syagriotermes salomeae Engel, Nel & Perrichot, 2011, мел (альб-сеноман), французский янтарь
- †Lebanotermes veltzae Engel, Azar & Nel, 2011, мел (апт), ливанский янтарь
Филогения
Молекулярно-генетический анализ подтвердил[127] давние взгляды, выдвинутые Кливлендом в 1934 году, о родстве термитов с субсоциальными древесными тараканами Cryptocercus[128]. Дополнительные данные были получены при анализе морфологических признаков нимф этих тараканов[129]. Эти сходства заставили некоторых авторов считать всех термитов единственным семейством Termitidae в рамках отряда тараканов Blattodea[6][130]. Другие исследователи настаивали на статусе термитов в ранге эписемейства Termitoidae, так как это позволит сохранить всю существующую классификацию термитов рангом ниже (семейства, подсемейства, трибы)[131]; или в ранге инфраотряда[3][4]. В некоторых современных энциклопедиях вообще избегают называть ранг группы термитов[132].
| Dictyoptera |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Классификация 2021 года
В 2021 году была пересмотрена классификация низших групп термитов в результате открытия новых ископаемых групп[125][133]. В 2024 году проведена реклассификация современных групп термитов Neoisoptera[123].
- Клада Xylophagodea Engel, 2011 (= Tutricablattae Evangelista et al., 2019)
- Инфраотряд Cryptocercaptera Engel, 2021 (диагноз эквивалентен Cryptocercidae)
- Семейство Cryptocercidae Handlirsch, 1925
- Инфраотряд Isoptera Brullé, 1832 (=Cratomastotermitidae + Mastotermitidae + Euisoptera)
- Семейство † Cratomastotermitidae Engel et al., 2009[5]
- †Cratomastotermes Bechly, 2007[134]
- †Cratomastotermes wolfschwenningeri Bechly, 2007 — меловой период, Бразилия
- †Cratomastotermes Bechly, 2007[134]
- Семейство Mastotermitidae Desneux, 1904
- Подсемейство †Idanotermitinae Engel, 2021
- †Idanotermes Engel, 2008[135]
- †Idanotermes desioculus Engel, 2008
- †Idanotermes Engel, 2008[135]
- Подсемейство Mastotermitinae Desneux, 1904
- Подсемейство †Idanotermitinae Engel, 2021
- Семейство † Cratomastotermitidae Engel et al., 2009[5]
- Инфраотряд Cryptocercaptera Engel, 2021 (диагноз эквивалентен Cryptocercidae)
- Парвотряд Euisoptera Engel et al., 2009 (4 семейства + Teletisoptera + Artisoptera)
- Семейство †Melqartitermitidae Engel, 2021
- †Melqartitermes Engel et al., 2007[136]
- †Melqartitermes myrrheus Engel et al., 2007 — ливанский янтарь
- †Melqartitermes Engel et al., 2007[136]
- Семейство †Mylacrotermitidae Engel, 2021
- †Mylacrotermes Engel et al., 2007[136]
- †Mylacrotermes cordatus Engel et al., 2007 — бирманский янтарь
- †Mylacrotermes Engel et al., 2007[136]
- Семейство †Krishnatermitidae Engel, 2021 (Возможно, это стволовая линия внутри Teletisoptera)
- †Krishnatermes Engel et al., 2016
- †Krishnatermes yoddha Engel et al. 2016 — бирманский янтарь
- †Krishnatermes Engel et al., 2016
- Семейство †Termopsidae Holmgren, 1911
- Семейство †Melqartitermitidae Engel, 2021
- Минотряд Teletisoptera Barden & Engel, 2021
- Семейство †Arceotermitidae Engel, 2021
- Подсемейство †Arceotermitinae Engel, 2021
- †Arceotermes Engel & Jiang, 2021
- †Arceotermes hospitis Engel & Jiang, 2021
- †Arceotermes Engel & Jiang, 2021
- Подсемейство †Cosmotermitinae Engel, 2021
- †Cosmotermes Zhao et al., 2020[137]
- Подсемейство †Arceotermitinae Engel, 2021
- Семейство Hodotermitidae Desneux, 1904
- Семейство Archotermopsidae Engel et al., 2009
- Подсемейство Hodotermopsinae Engel, 2021 → Hodotermopsidae (2024)
- Подсемейство Archotermopsinae Engel et al., 2009
- Семейство Stolotermitidae Holmgren, 1910 (В некоторых исследованиях это семейство относят к Artisoptera как сестринское к Icoisoptera (Engel et al., 2009, 2016; Jouault et al., 2021))[5][138][139]
- Подсемейство Stolotermitinae Holmgren, 1910
- Подсемейство Porotermitinae Emerson, 1942
- Семейство †Arceotermitidae Engel, 2021
- Минотряд Artisoptera Engel, 2021 (=Tanytermitidae + Icoisoptera)
- Семейство †Tanytermitidae Engel, 2021
- †Tanytermes Engel et al., 2007
- †Tanytermes anawrahtai Engel et al. 2007 — бирманский янтарь
- †Tanytermitalis Engel & Cai, 2021
- †Tanytermitalis philetaerus Engel & Cai, 2021
- †Tanytermes Engel et al., 2007
- Семейство †Tanytermitidae Engel, 2021
- Microrder Icoisoptera Engel in Engel et al., 2013 (=Kalotermitidae + Neoisoptera)
- Семейство Kalotermitidae Froggatt, 1897
- Nanorder Neoisoptera Engel et al., 2009 (высшие термиты)
Neoisoptera
Клада или нанотряд Neoisoptera, также именуемая «новейшие термиты» (в эволюционном смысле), включает так называемых «высших термитов», хотя некоторые авторы применяют этот термин только к крупнейшему семейству Termitidae. Для последних характерно отсутствие нимф псевдоэргат (многие нимфы рабочих «низших термитов» обладают способностью развиваться в репродуктивные касты). Переваривание целлюлозы у «высших термитов» эволюционировало совместно с кишечной эукариотической микробиотой[140] и многие роды имеют симбиотические отношения с грибами, такими как Termitomyces; напротив, «низшие термиты» обычно содержат жгутиковых и прокариот в заднем кишечнике. В 2024 году проведена реклассификация современных групп термитов Neoisoptera и дополнительно выделены семейства Heterotermitidae, Psammotermitidae и Termitogetonidae (признано 13 современных семейств термитов и 8 вымерших)[123]. Cемейства высших термитов:
|
|
См. также
- Mastotermes darwiniensis
- Туркменский термит
Примечания
- ↑ Жизнь животных. В 7 т. / гл. ред. В. Е. Соколов. — 2‑е изд., перераб. — М. : Просвещение, 1984. — Т. 3 : Членистоногие: трилобиты, хелицеровые, трахейнодышащие. Онихофоры / под ред. М. С. Гилярова, Ф. Н. Правдина. — С. 166. — 463 с. : ил.
- ↑ Белые муравьи Архивная копия от 26 ноября 2020 на Wayback Machine. БСЭ (1-е издание). Москва, 1926—1947. Том. V (1927): Барыкова — Бессалько, стлб. 442.
- ↑ 1 2 3 Engel Michael S. Family-group names for termites (Isoptera), redux (англ.) // ZooKeys. — Pensoft Publishers, 2011. — Vol. 148. — P. 171—184. Архивировано 22 октября 2012 года.
- ↑ 1 2 3 4 5 Krishna, Kumar; Grimaldi, David A.; Krishna, Valerie.; Engel, Michael S. Treatise on the Isoptera of the world (англ.) // Bulletin of the American Museum of Natural History[англ.] : Журнал. — Нью-Йорк: American Museum of Natural History, 2013. — Vol. 377 (vol. 1-7). — P. 1—2704. — ISSN 0003-0090. Архивировано 30 июля 2014 года.
- ↑ 1 2 3 4 5 Engel M. S., Grimaldi D. A., Krishna K. 2009. Termites (Isoptera): Their Phylogeny, Classification, and Rise to Ecological Dominance Архивная копия от 28 июля 2009 на Wayback Machine. American Museum Novitates 3650:1—27.
- ↑ 1 2 Inward, D., G. Beccaloni, and P. Eggleton. Death of an order: a comprehensive molecular phylogenetic study confirms that termites are eusocial cockroaches (англ.) // Biology Letters. — Royal Society Publishing, 2007. — Vol. 3, no. 3. — P. 331—335. — doi:10.1098/rsbl.2007.0367.
- ↑ Dominic A. Evangelista, Benjamin Wipfler, Olivier Béthoux, Alexander Donath, Mari Fujita, Manpreet K. Kohli, Frédéric Legendre, Shanlin Liu, Ryuichiro Machida, Bernhard Misof, Ralph S. Peters, Lars Podsiadlowski, Jes Rust, Kai Schuette, Ward Tollenaar, Jessica L. Ware, Torsten Wappler, Xin Zhou, Karen Meusemann & Sabrina Simon. An integrative phylogenomic approach illuminates the evolutionary history of cockroaches and termites (Blattodea) (англ.) // Proceedings of the Royal Society B : Журнал. — Royal Society, 2019. — Vol. 286, no. 1895. — P. 1—9. — ISSN 1471-2954. — doi:10.1098/rspb.2018.2076. Архивировано 11 апреля 2021 года.
- ↑ Bignell, 2010.
- ↑ Thorne, Barbara L. Evolution of eusociality in termites (англ.) // Annual Review of Ecology, Evolution, and Systematics : journal. — Annual Reviews, 1997. — Vol. 28, no. 5. — P. 27—53. — doi:10.1146/annurev.ecolsys.28.1.27. — PMC 349550. Архивировано 30 мая 2010 года.
- ↑ Эдвард Уилсон. Эусоциальность. Люди, муравьи, голые землекопы и другие общественные животные = Wilson Edward. Genesis: The Deep Origin of Societies. — М.: Альпина нон-фикшн, 2019. — С. 62. — 158 с. — ISBN 978-5-91671-697-9.
- ↑ 1 2 Rudolf H. Scheffrahn. Termites (Isoptera) (англ.) / Ed. John L. Capinera. — Encyclopedia of Entomology. — Springer Netherlands, 2008. — Vol. 20. — P. 3737—3747. — ISBN 978-1-4020-6242-1. — doi:10.1007/978-1-4020-6359-6_2400.
- ↑ Christine A. Nalepa. Body Size and Termite Evolution (англ.) // Evolutionary Biology : Журнал. — 2011. — Vol. 38, no. 3. — P. 243—257. (недоступная ссылка)
- ↑ Insect Biodiversity: science and society / Robert G. Foottit, Peter H. Adler.. — Blackwell Publishing Ltd, 2009. — С. 31. — 642 с. — ISBN 978-1-4051-5142-9. Архивировано 3 апреля 2018 года.
- ↑ Marie Djernæs. Biodiversity of Blattodea – the Cockroaches and Termites (Chapter 14) // Insect Biodiversity: Science and Society, II / Robert G. Foottit, Peter H. Adler.. — First Edition. — John Wiley & Sons Ltd, 2018. — С. 359—387. — 987 с. — ISBN 978-1-1189-4558-2. Архивировано 24 декабря 2019 года.
- ↑ В Узбекистане термитами заражено более 25 тысяч жилых объектов. Regnum (25 апреля 2009). Дата обращения: 14 августа 2010. Архивировано 29 апреля 2009 года.
- ↑ 1 2 3 4 Termite Biology and Ecology. Division of Technology, Industry and Economics Chemicals Branch. United Nations Environment Programme. Дата обращения: 12 января 2015. Архивировано из оригинала 10 ноября 2014 года.
- ↑ Meyer, V.W. Distribution and density of termite mounds in the northern Kruger National Park, with specific reference to those constructed by Macrotermes Holmgren (Isoptera: Termitidae) (англ.) // African Entomology : journal. — 1999. — Vol. 7, no. 1. — P. 123—130. Архивировано 4 октября 2018 года.
- ↑ Sanderson, M.G. Biomass of termites and their emissions of methane and carbon dioxide: A global database (англ.) // Global Biogeochemical Cycles : journal. — 1996. — Vol. 10, no. 4. — P. 543—557. — doi:10.1029/96GB01893. — .
- ↑ Heather, N.W. The exotic drywood termite Cryptotermes brevis (Walker) (Isoptera : Kalotermitidae) and endemic Australian drywood termites in Queensland (англ.) // Australian Journal of Entomology : journal. — Wiley-Blackwell, 1971. — Vol. 10, no. 2. — P. 134—141. — doi:10.1111/j.1440-6055.1971.tb00022.x.
- ↑ 1 2 3 4 5 6 Termites: evolution, sociality, symbioses, ecology / Ed. by Takuya Abe (01.03.1945-27.03.2000), David Edward Bignell, Masahiko Higashi (11.08.1954-27.03.2000 ). — Dordrecht, Netherlands: Kluwer Academic Publishers, 2000. — 467 с. — ISBN 0-7923-6361-2. Архивировано 5 ноября 2015 года.
- ↑ Thomas E. Snyder. (1949). Catalog of termites (Isoptera) of the world. Архивная копия от 11 сентября 2016 на Wayback Machine — Smithsonian miscellaneous collections, v. 112. — Washington, Smithsonian Institution, 1949. — 490pp.
- ↑ Элементы — новости науки: Кастовая принадлежность у термитов предопределена генетически. Дата обращения: 24 ноября 2007. Архивировано 17 ноября 2007 года.
- ↑ Some female termites can reproduce without males Архивная копия от 30 сентября 2018 на Wayback Machine, September 25, 2018
- ↑ Kenji Matsuura. Sexual and Asexual Reproduction in Termites / Ed. David Edward Bignell, Yves Roisin, Nathan Lo. — Biology of Termites: a Modern Synthesis. — Springer Netherlands, 2011. — С. 255—277. — ISBN 978-90-481-3976-7. (недоступная ссылка)
- ↑ Silvia Bergamaschi, Tracy Z. Dawes-Gromadzki, Valerio Scali, Mario Marini and Barbara Mantovani. 2007. Karyology, mitochondrial DNA and the phylogeny of Australian termites Архивная копия от 15 сентября 2022 на Wayback Machine. — Chromosome Research. Volume 15, Number 6 (2007), 735—753.
- ↑ Blackmon, Heath. Synthesis and phylogenetic comparative analyses of the causes and consequences of karyotype evolution in arthropods (англ.). — Arlington: The University of Texas at Arlington, 2015. — 447 p. — (Degree of Doctor of Philosophy). Архивировано 14 сентября 2022 года.
- ↑ Mizumoto, Nobuaki; Bourguignon, Thomas; Bailey, Nathan W. (2022-11-15). "Ancestral sex-role plasticity facilitates the evolution of same-sex sexual behavior". Proceedings of the National Academy of Sciences (англ.). 119 (46): e2212401119. Bibcode:2022PNAS..11912401M. doi:10.1073/pnas.2212401119. ISSN 0027-8424. PMC 9674213. PMID 36346843.
- ↑ Eisuke Tasaki, Kazuya Kobayashi, Kenji Matsuura, Yoshihito Iuchi. (2017). An Efficient Antioxidant System in a Long-Lived Termite Queen Архивная копия от 20 января 2017 на Wayback Machine. — PLoS ONE 12(1): e0167412. https://dx.doi.org/10.1371/journal.pone.0167412
- ↑ Bordereau, C.; A. Robert, V. Van Tuyen and A. Peppuy. Suicidal defensive behaviour by frontal gland dehiscence in Globitermes sulphureus Haviland soldiers (Isoptera) (англ.) // Insectes Sociaux : journal. — Birkhäuser Basel, 1997. — August (vol. 44, no. 3). — P. 289—297. — ISSN 1420-9098. Архивировано 18 апреля 2020 года.
- ↑ 1 2 Henry R. Hermann (ed.). (1981). Social Insects Архивная копия от 28 августа 2016 на Wayback Machine. Volume 2. New York: Academic Press, 1981. xiii + pp.1-502. ISBN 0-12-342202-7. — Jean Deligne, Andre Quennedey, Murray S. Blum. The Enemies ad Defense Mechanisms of Termites. Pp. 2-76.
- ↑ Ahmad M. (1976). The soldierlles termite genera of the Oriental region, with a note on their phylogeny (Isoptera Termitidae). Pakistan Journal ol Zoology 8, 105—123.
- ↑ Miller L. R. (1984). Invasitermes, a new genus of soldierlles termites from Northern Australia (Isoptera Termitidae). Journal of the Australian Entomological Society 23: 33-37
- ↑ Bignell, Roisin, Lo, 2010, p. 11.
- ↑ Traniello J. F. A., Leuthold R. H. Behavior and Ecology of Foraging in Termites // Termites : evolution, sociality, symbioses, ecology. — Dordrecht: Springer Netherlands, 2000. — С. 141–168. — 466 с. — ISBN 978-94-017-3223-9, 94-017-3223-X. — doi:10.1007/978-94-017-3223-9_7. Архивировано 27 января 2022 года.
- ↑ Bignell, Roisin, Lo, 2010, p. 13.
- ↑ Reinhard J., Kaib M. Trail Communication During Foraging and Recruitment in the Subterranean Termite Reticulitermes santonensis De Feytaud (Isoptera, Rhinotermitidae) // Journal of Insect Behavior. — 2004. — doi:10.1023/A:1007881510237. Архивировано 26 января 2022 года.
- ↑ Costa-Leonardo A. M., Haifig I. Termite Communication During Different Behavioral Activities (англ.) // Biocommunication of Animals / Guenther Witzany. — Dordrecht: Springer Netherlands, 2014. — P. 161–190. — ISBN 978-94-007-7414-8. — doi:10.1007/978-94-007-7414-8_10.
- ↑ Costa-Leonardo A. M. Morphology of the sternal gland in workers of Coptotermes gestroi (Isoptera, Rhinotermitidae) (англ.) // Micron. — 2006-08-01. — Vol. 37, iss. 6. — P. 551–556. — ISSN 0968-4328. — doi:10.1016/j.micron.2005.12.006.
- ↑ Traniello J. F., Busher C. Chemical regulation of polyethism during foraging in the neotropical termite Nasutitermes costalis (англ.) // Journal of Chemical Ecology. — 1985-03-01. — Vol. 11, iss. 3. — P. 319–332. — ISSN 1573-1561. — doi:10.1007/BF01411418.
- ↑ Miramontes O., DeSouza O., Paiva L. R., Marins A., Orozco S., Aegerter C. M. Lévy Flights and Self-Similar Exploratory Behaviour of Termite Workers: Beyond Model Fitting // PLoS ONE. — 2014-10-29. — Т. 9, вып. 10. — С. e111183. — ISSN 1932-6203. — doi:10.1371/journal.pone.0111183. Архивировано 10 июля 2022 года.
- ↑ 1 2 Mizumoto N., Bourguignon T., Kanao T. Termite nest evolution fostered social parasitism by termitophilous rove beetles (англ.) // Evolution. — 2022. — Vol. 76(5). — P. 1064—1072. — doi:10.1111/evo.14457. — PMID 35319096.
- ↑ Jost, C.; Haifig, I.; de Camargo-Dietrich, C.R.R.; Costa-Leonardo, A.M. A comparative tunnelling network approach to assess interspecific competition effects in termites (англ.) // Insectes Sociaux : journal. — 2012. — Vol. 59, no. 3. — P. 369—379. — doi:10.1007/s00040-012-0229-7.
- ↑ Polizzi, J.M.; Forschler, B.T. Intra- and interspecific agonism in Reticulitermes flavipes (Kollar) and R. virginicus (Banks) and effects of arena and group size in laboratory assays (англ.) // Insectes Sociaux : journal. — 1998. — Vol. 45, no. 1. — P. 43—49. — doi:10.1007/s000400050067.
- ↑ Darlington, J.P.E.C. The underground passages and storage pits used in foraging by a nest of the termite Macrotermes michaelseni in Kajiado, Kenya (англ.) // Journal of Zoology : journal. — Wiley-Blackwell, 1982. — Vol. 198, no. 2. — P. 237—247. — doi:10.1111/j.1469-7998.1982.tb02073.x.
- ↑ 1 2 3 4 5 6 7 8 9 Costa-Leonardo, A.M.; Haifig, I. Termite communication dduring different behavioral activities in Biocommunication of Animals (англ.). — Springer Netherlands, 2013. — P. 161—190. — ISBN 978-94-007-7413-1. — doi:10.1007/978-94-007-7414-8_10.
- ↑ Cornelius, M.L.; Osbrink, W.L. Effect of soil type and moisture availability on the foraging behavior of the Formosan subterranean termite (Isoptera: Rhinotermitidae) (англ.) // Journal of Economic Entomology : journal. — 2010. — Vol. 103, no. 3. — P. 799—807. — doi:10.1603/EC09250. — PMID 20568626.
- ↑ Toledo Lima, J.; Costa-Leonardo, A.M. Subterranean termites (Isoptera: Rhinotermitidae): Exploitation of equivalent food resources with different forms of placement (англ.) // Insect Science : journal. — 2012. — Vol. 19, no. 3. — P. 412—418. — doi:10.1111/j.1744-7917.2011.01453.x.
- ↑ Jmhasly, P.; Leuthold, R.H. Intraspecific colony recognition in the termites Macrotermes subhyalinus and Macrotermes bellicosus (Isoptera, Termitidae) (англ.) // Insectes Sociaux : journal. — 1999. — Vol. 46, no. 2. — P. 164—170. — doi:10.1007/s000400050128.
- ↑ Messenger, M.T.; Su, N.Y. Agonistic behavior between colonies of the Formosan subterranean termite (Isoptera: Rhinotermitidae) from Louis Armstrong Park, New Orleans, Louisiana (англ.) // Sociobiology : journal. — 2005. — Vol. 45, no. 2. — P. 331—345.
- ↑ Korb, J.; Weil, T.; Hoffmann, K.; Foster, K.R.; Rehli, M. A gene necessary for reproductive suppression in termites (англ.) // Science : journal. — 2009. — Vol. 324, no. 5928. — P. 758. — doi:10.1126/science.1170660. — . — PMID 19423819.
- ↑ Mathew, T.T.G.; Reis, R.; DeSouza, O.; Ribeiro, S.P. Predation and interference competition between ants (Hymenoptera: Formicidae) and arboreal termites (Isoptera: Termitidae) (англ.) // Sociobiology : journal. — 2005. — Vol. 46, no. 2. — P. 409—419. Архивировано 11 апреля 2021 года.
- ↑ 1 2 3 Costa-Leonardo, A.M.; Haifig, I. Pheromones and exocrine glands in Isoptera // Vitamins and Hormones. — 2010. — Т. 83. — С. 521—549. — doi:10.1016/S0083-6729(10)83021-3. — PMID 20831960.
- ↑ 1 2 3 Krishna, K. Termite. Encyclopædia Britannica. Дата обращения: 11 сентября 2015. Архивировано 21 апреля 2021 года.
- ↑ Evans, T.A.; Inta, R.; Lai, J.C.S.; Lenz, M. Foraging vibration signals attract foragers and identify food size in the drywood termite, Cryptotermes secundus (англ.) // Insectes Sociaux : journal. — 2007. — Vol. 54, no. 4. — P. 374—382. — doi:10.1007/s00040-007-0958-1.
- ↑ Costa-Leonardo, A.M.; Casarin, F.E.; Lima, J.T. Chemical communication in isoptera // Neotropical Entomology. — 2009. — Т. 38, № 1. — С. 1—6. — doi:10.1590/S1519-566X2009000100001. — PMID 19347093.
- ↑ Richard, F.-J.; Hunt, J.H. Intracolony chemical communication in social insects (англ.) // Insectes Sociaux : journal. — 2013. — Vol. 60, no. 3. — P. 275—291. — doi:10.1007/s00040-013-0306-6. Архивировано 4 марта 2016 года.
- ↑ Dronnet, S.; Lohou, C.; Christides, J.P.; Bagnères, A.G. Cuticular hydrocarbon composition reflects genetic relationship among colonies of the introduced termite Reticulitermes santonensis Feytaud (англ.) // Journal of Chemical Ecology[англ.] : journal. — Springer, 2006. — Vol. 32, no. 5. — P. 1027—1042. — doi:10.1007/s10886-006-9043-x. — PMID 16739021.
- ↑ Rosengaus R. B., Traniello J. F. A., Chen T., Brown J. J., Karp R. D. Immunity in a social insect // Naturwissenschaften. — 1999. — Vol. 86. — P. 588–591. — doi:10.1007/s001140050679. — .
- ↑ Wilson D. S. Above ground predator defense in the harvester termite, Hodotermes mossambicus (Hagen) // Journal of the Entomological Society of Southern Africa. — 1977. — Vol. 40. — P. 271–282.
- ↑ Matsuura K. Termite-egg mimicry by a sclerotium-forming fungus // Proceedings of the Royal Society B: Biological Sciences. — 2006. — Vol. 273. — P. 1203–1209. — doi:10.1098/rspb.2005.3434. — PMID 16720392. — PMC 1560272.
- ↑ Matsuura K., Yashiro T., Shimizu K., Tatsumi S., Tamura T. Cuckoo fungus mimics termite eggs by producing the cellulose-digesting enzyme β-glucosidase // Current Biology. — 2009. — Vol. 19. — P. 30–36. — doi:10.1016/j.cub.2008.11.030. — PMID 19110429.
- ↑ Howard R. W., McDaniel C. A., Blomquist G. J. Chemical mimicry as an integrating mechanism: cuticular hydrocarbons of a termitophile and its host // Science. — 1980. — Vol. 210. — P. 431–433. — doi:10.1126/science.210.4468.431. — . — PMID 17837424.
- ↑ Watson J. A. L. Austrospirachtha mimetes a new termitophilous corotocine from Northern Australia (Coleoptera: Staphylinidae) // Australian Journal of Entomology. — 1973. — Vol. 12. — P. 307–310. — doi:10.1111/j.1440-6055.1973.tb01678.x.
- ↑ 1 2 3 Bignell, 2010, p. 21.
- ↑ 1 2 Bignell, 2010, p. 22.
- ↑ Perna, A.; Jost, C.; Couturier, E.; Valverde, S.; Douady, S.; Theraulaz, G. The structure of gallery networks in the nests of termite Cubitermes spp. revealed by X-ray tomography (англ.) // Die Naturwissenschaften : journal. — 2008. — Vol. 95, no. 9. — P. 877—884. — doi:10.1007/s00114-008-0388-6. — . — PMID 18493731.
- ↑ Glenday, Craig. Guinness World Records 2014. — 2014. — С. 33. — ISBN 978-1-908843-15-9.
- ↑ Jacklyn, P. Evidence for Adaptive Variation in the Orientation of Amitermes (Isoptera, Termitinae) Mounds From Northern Australia (англ.) // Australian Journal of Zoology : journal. — 1991. — Vol. 39, no. 5. — P. 569. — doi:10.1071/ZO9910569.
- ↑ Jacklyn, P.M.; Munro, U. Evidence for the use of magnetic cues in mound construction by the termite Amitermes meridionalis (Isoptera : Termitinae) (англ.) // Australian Journal of Zoology : journal. — 2002. — Vol. 50, no. 4. — P. 357. — doi:10.1071/ZO01061.
- ↑ Grigg, G.C. Some Consequences of the Shape and Orientation of 'magnetic' Termite Mounds (англ.) // Australian Journal of Zoology : journal. — 1973. — Vol. 21, no. 2. — P. 231—237. — doi:10.1071/ZO9730231.
- ↑ Kok O. B., Hewitt P. H. Bird and mammal predators of the harvester termite Hodotermes mossambicus (Hagen) in semi-arid regions of South Africa (англ.) // South African Journal of Science. — 1990. — Vol. 86, no. 1. — P. 34—37. — ISSN 0038-2353.
- ↑ 1 2 3 4 5 Hölldobler B., Wilson E. O. The Ants. — Cambridge, Massachusetts: Harvard University Press, 1990. — С. 559—566. — ISBN 978-0-674-04075-5.
- ↑ 1 2 Culliney, T.W.; Grace, J.K. Prospects for the biological control of subterranean termites (Isoptera: Rhinotermitidae), with special reference to Coptotermes formosanus (англ.) // Bulletin of Entomological Research : journal. — 2000. — Vol. 90, no. 1. — P. 9—21. — doi:10.1017/S0007485300000663. — PMID 10948359.
- ↑ Reagan, D.P.; Waide, R.B. The food web of a tropical rain forest. — Chicago: University of Chicago Press, 1996. — С. 294. — ISBN 978-0-226-70599-6.
- ↑ Wade, W.W. Ecology of Desert Systems. — Burlington: Elsevier, 2002. — С. 216. — ISBN 978-0-08-050499-5.
- ↑ Dean, W.R.J.; Milton, S.J. Plant and invertebrate assemblages on old fields in the arid southern Karoo, South Africa (англ.) // African Journal of Ecology. — 1995. — Vol. 33, no. 1. — P. 1—13. — doi:10.1111/j.1365-2028.1995.tb00777.x.
- ↑ 1 2 Bardgett, R.D.; Herrick, J.E.; Six, J.; Jones, T.H.; Strong, D.R.; van der Putten, W.H. Soil ecology and ecosystem services (англ.). — 1st. — Oxford: Oxford University Press, 2013. — P. 178. — ISBN 978-0-19-968816-6.
- ↑ Bignell, 2010, p. 509.
- ↑ Choe, J.C.; Crespi, B.J. The evolution of social behavior in insects and arachnids (англ.). — 1st. — Cambridge: Cambridge university press, 1997. — P. 76. — ISBN 978-0-521-58977-2.
- ↑ 1 2 Abe Y., Bignell D.E., Higashi T. Termites: Evolution, Sociality, Symbioses, Ecology (англ.). — Springer, 2014. — P. 124—149. — ISBN 978-94-017-3223-9. — doi:10.1007/978-94-017-3223-9.
- ↑ Wilson D.S., Clark A.B. Above ground defence in the harvester termite, Hodotermes mossambicus (англ.) // Journal of the Entomological Society of South Africa. — 1977. — Vol. 40. — P. 271—282.
- ↑ Lavelle, P.; Spain, A.V. Soil ecology. — 2nd. — Dordrecht: Kluwer Academic, 2001. — С. 316. — ISBN 978-0-306-48162-8.
- ↑ Richardson, P.K.R.; Bearder, S.K. (1984). "The Hyena Family". In MacDonald, D. (ed.). The Encyclopedia of Mammals. New York, NY: Facts on File Publication. pp. 158—159. ISBN 978-0-87196-871-5.
- ↑ Mills, G.; Harvey, M. African Predators. — Washington, D.C.: Smithsonian Institution Press, 2001. — С. 71. — ISBN 978-1-56098-096-4.
- ↑ d'Errico F., Backwell L. Assessing the function of early hominin bone tools (англ.) // Journal of Archaeological Science[англ.]. — Elsevier, 2009. — Vol. 36, no. 8. — P. 1764—1773. — doi:10.1016/j.jas.2009.04.005.
- ↑ Lepage, M.G. Étude de la prédation de Megaponera foetens (F.) sur les populations récoltantes de Macrotermitinae dans un ecosystème semi-aride (Kajiado-Kenya) (исп.) // Insectes Sociaux. — 1981. — V. 28, no 3. — P. 247—262. — doi:10.1007/BF02223627.
- ↑ Levieux, J. Note préliminaire sur les colonnes de chasse de Megaponera fœtens F. (Hyménoptère Formicidæ) (фр.) // Insectes Sociaux. — 1966. — Vol. 13, no 2. — P. 117—126. — doi:10.1007/BF02223567.
- ↑ Longhurst, C.; Baker, R.; Howse, P.E. Chemical crypsis in predatory ants // Experientia. — 1979. — Т. 35, № 7. — С. 870—872. — doi:10.1007/BF01955119.
- ↑ Wheeler W.M. Ecological relations of Ponerine and other ants to termites (англ.) // Proceedings of the American Academy of Arts and Sciences : journal. — 1936. — Vol. 71, no. 3. — P. 159—171. — doi:10.2307/20023221. — .
- ↑ Shattuck, S.O.; Heterick, B.E. Revision of the ant genus Iridomyrmex (Hymenoptera : Formicidae) (англ.) // Zootaxa : journal. — 2011. — Vol. 2845. — P. 1—74. — ISBN 978-1-86977-676-3. — ISSN 1175-5334. Архивировано 15 марта 2021 года.
- ↑ Traniello, J.F.A. Enemy deterrence in the recruitment strategy of a termite: Soldier-organized foraging in Nasutitermes costalis (англ.) // Proceedings of the National Academy of Sciences : journal. — National Academy of Sciences, 1981. — Vol. 78, no. 3. — P. 1976—1979. — doi:10.1073/pnas.78.3.1976. — . — PMID 16592995. — PMC 319259.
- ↑ Schöning C., Moffett M.W. Driver Ants Invading a Termite Nest: why do the most catholic predators of all seldom take this abundant prey? (англ.) // Biotropica[англ.] : journal. — Wiley-Blackwell, 2007. — Vol. 39, no. 5. — P. 663—667. — doi:10.1111/j.1744-7429.2007.00296.x. Архивировано 12 ноября 2015 года.
- ↑ Mill, A.E. Observations on Brazilian termite alate swarms and some structures used in the dispersal of reproductives (Isoptera: Termitidae) (англ.) // Journal of Natural History : journal. — Taylor & Francis, 1983. — Vol. 17, no. 3. — P. 309—320. — doi:10.1080/00222938300770231.
- ↑ Schmid-Hempel, 1998, p. 61.
- ↑ Schmid-Hempel, 1998, p. 75.
- ↑ Wilson, E.O. The Insect Societies. — 5th. — Cambridge, Massachusetts: Harvard University Press, 1971. — Т. 76. — С. 398. — ISBN 978-0-674-45495-8.
- ↑ Schmid-Hempel, 1998, p. 59.
- ↑ Schmid-Hempel, 1998, pp. 301–302.
- ↑ Schmid-Hempel, 1998, p. 19.
- ↑ Weiser, J.; Hrdy, I. Pyemotes – mites as parasites of termites // Zeitschrift für Angewandte Entomologie. — 2009. — Т. 51, № 1—4. — С. 94—97. — doi:10.1111/j.1439-0418.1962.tb04062.x.
- ↑ 1 2 Chouvenc, T.; Efstathion, C.A.; Elliott, M.L.; Su, N.Y. Resource competition between two fungal parasites in subterranean termites (англ.) // Die Naturwissenschaften : journal. — 2012. — Vol. 99, no. 11. — P. 949—958. — doi:10.1007/s00114-012-0977-2. — . — PMID 23086391.
- ↑ Schmid-Hempel, 1998, pp. 38, 102.
- ↑ Schmid-Hempel, 1998, p. 116.
- ↑ Corinne Rouland-Lefèvre. Termites as Pests of Agriculture / Ed. David Edward Bignell, Yves Roisin, Nathan Lo. — Biology of Termites: a Modern Synthesis. — Springer Netherlands, 2011. — С. 499—517. — ISBN 978-90-481-3976-7. (недоступная ссылка)
- ↑ Ye Weimin et al. (2004). Phylogenetic relationships of nearctic Reticulitermes species (Isoptera: Rhinotermitidae) with particular reference to Reticulitermes arenincola Goellner. — Molecular Phylogenetics and Evolution. — 30(2004): 815—822.
- ↑ «Гумбольдт, возвратясь из путешествия по Южной Америке, с сожалением отмечал, что на этом континенте редко увидишь книгу старше пятидесяти лет». Акимушкин И. «Мир животных. Рассказы о насекомых», Москва, «Молодая гвардия», 1975
- ↑ Theodore A. Evans. Invasive Termites / Ed. David Edward Bignell, Yves Roisin, Nathan Lo. — Biology of Termites: a Modern Synthesis. — 2011. — С. 519—562. — ISBN 978-90-481-3976-7. (недоступная ссылка)
- ↑ 1 2 Figueirêdo, R.E.C.R.; Vasconcellos, A.; Policarpo, I.S.; Alves, R.R.N. Edible and medicinal termites: a global overview // Journal of Ethnobiology and Ethnomedicine. — 2015. — Т. 11, № 1. — С. 1—17. — doi:10.1186/s13002-015-0016-4. — PMID 25925503. — PMC 4427943.
- ↑ 1 2 3 4 Nyakupfuka, A. Global Delicacies: Discover Missing Links from Ancient Hawaiian Teachings to Clean the Plaque of your Soul and Reach Your Higher Self (англ.). — Bloomington, Indiana: BalboaPress, 2013. — P. 40—41. — ISBN 978-1-4525-6791-4.
- ↑ 1 2 Bodenheimer, F.S. Insects as Human Food: A Chapter of the Ecology of Man (англ.). — Netherlands: Springer, 1951. — P. 331—350. — ISBN 978-94-017-6159-8.
- ↑ Geissler, P.W. The significance of earth-eating: social and cultural aspects of geophagy among Luo children (англ.) // Africa : journal. — 2011. — Vol. 70, no. 4. — P. 653—682. — doi:10.3366/afr.2000.70.4.653.
- ↑ Knudsen, J.W. Akula udongo (earth eating habit): a social and cultural practice among Chagga women on the slopes of Mount Kilimanjaro (англ.) // African Journal of Indigenous Knowledge Systems : journal. — 2002. — Vol. 1, no. 1. — P. 19—26. — ISSN 1683-0296. — doi:10.4314/indilinga.v1i1.26322.
- ↑ Nchito, M.; Wenzel Geissler, P.; Mubila, L.; Friis, H.; Olsen, A. Effects of iron and multimicronutrient supplementation on geophagy: a two-by-two factorial study among Zambian schoolchildren in Lusaka (англ.) // Transactions of the Royal Society of Tropical Medicine and Hygiene : journal. — 2004. — Vol. 98, no. 4. — P. 218—227. — doi:10.1016/S0035-9203(03)00045-2. — PMID 15049460.
- ↑ Saathoff, E.; Olsen, A.; Kvalsvig, J.D.; Geissler, P.W. Geophagy and its association with geohelminth infection in rural schoolchildren from northern KwaZulu-Natal, South Africa (англ.) // Transactions of the Royal Society of Tropical Medicine and Hygiene : journal. — 2002. — Vol. 96, no. 5. — P. 485—490. — doi:10.1016/S0035-9203(02)90413-X. — PMID 12474473.
- ↑ Katayama, N.; Ishikawa, Y.; Takaoki, M.; Yamashita, M.; Nakayama, S.; Kiguchi, K.; Kok, R.; Wada, H.; Mitsuhashi, J. Entomophagy: A key to space agriculture (англ.) // Advances in Space Research[англ.]. — Elsevier, 2008. — Vol. 41, no. 5. — P. 701—705. — doi:10.1016/j.asr.2007.01.027. — . Архивировано 27 сентября 2018 года.
- ↑ Жужиков, 1979, с. 7.
- ↑ Метерлинк М. Тайная жизнь термитов. — М.: Эксмо-Пресс, 2002. — 400 с.
- ↑ Theodore A. Evans, Tracy Z. Dawes, Philip R. Ward & Nathan Lo. Ants and termites increase crop yield in a dry climate (англ.) // Nature Communications : Журнал. — London: Nature Publishing Group, 2011. — Vol. 2, no. 262. — ISSN 2041-1723. Архивировано 22 февраля 2014 года. doi:10.1038/ncomms1257
- ↑ Результаты для отряда Isoptera на https://www.iucnredlist.org/search Архивная копия от 28 декабря 2021 на Wayback Machine (Дата обращения: 16 августа 2020)
- ↑ Gerlach J. 2012. Glyptotermes scotti Архивная копия от 2 июня 2018 на Wayback Machine. The IUCN Red List of Threatened Species 2012.
- ↑ Gerlach J. 2012. Procryptotermes fryeri Архивная копия от 2 июня 2018 на Wayback Machine. The IUCN Red List of Threatened Species 2012.
- ↑ Gerlach J. 2012. Nasutitermes maheensis Архивная копия от 18 сентября 2020 на Wayback Machine. The IUCN Red List of Threatened Species 2012.
- ↑ 1 2 3 4 Hellemans, S., Rocha, M.M., Wang, M. et al. Genomic data provide insights into the classification of extant termites (англ.) // Nature Communications : Журнал. — Великобритания: Nature Portfolio, 2024. — Vol. 15, no. 6724. — P. 1—17 (suppl.1—4: 1—50). — ISSN 2041-1723. — doi:10.1038/s41467-024-51028-y. Архивировано 9 августа 2024 года. Published: 07 August 2024
- ↑ Жужиков, 1979, с. 20.
- ↑ 1 2 Jiang R. X.; Zhang H. R.; Eldredge K. T.; Song X. B.; Li Y. D.; Tihelka E.; Huang D. Y.; Wang S.; Engel, M.S.; Cai C. Y. Further evidence of Cretaceous termitophily: Description of new termite hosts of the trichopseniine Cretotrichopsenius (Coleoptera: Staphylinidae), with emendations to the classification of lower termites (Isoptera) (англ.) // Palaeoentomology : журнал. — 2021. — Vol. 4, no. 4. — P. 374—389. — ISSN 2624-2826. — doi:10.11646/palaeoentomology.4.4.13.
- ↑ Zhipeng Zhao, Paul Eggleton, Xiangchu Yin, Taiping Gao, Chungkun Shih. The oldest known mastotermitids (Blattodea: Termitoidae) and phylogeny of basal termites (англ.) // Systematic Entomology. — Wiley-Blackwell, 2019. — Vol. 44, iss. 3. — P. 612—623. — ISSN 1365-3113. — doi:10.1111/syen.12344. Архивировано 14 сентября 2020 года.
- ↑ Ware, J.L.; Litman, J.; Klass, K.-D.; Spearman, L.A. Relationships among the major lineages of Dictyoptera: the effect of outgroup selection on dictyopteran tree topology (англ.) // Systematic Entomology : journal. — Wiley-Blackwell, 2008. — Vol. 33, no. 3. — P. 429—450. — doi:10.1111/j.1365-3113.2008.00424.x.
- ↑ Cleveland, L.R.; Hall, S.K.; Sanders, E.P.; Collier, J. The Wood-Feeding Roach Cryptocercus, its protozoa, and the symbiosis between protozoa and roach (англ.) // Memoirs of the American Academy of Arts and Sciences : journal. — 1934. — Vol. 17, no. 2. — P. 185—382. — doi:10.1093/aesa/28.2.216.
- ↑ McKittrick, F.A. A contribution to the understanding of cockroach-termite affinities (англ.) // Annals of the Entomological Society of America : journal. — 1965. — Vol. 58, no. 1. — P. 18—22. — doi:10.1093/aesa/58.1.18. — PMID 5834489.
- ↑ Eggleton P., Beccaloni G., Inward D. Response to Lo et al. (англ.) // Biology Letters. — Royal Society Publishing, 2007. — Vol. 3, no. 5. — P. 564—565.
- ↑ Lo N., Engel M.S., Cameron S., Nalepa C.A., Tokuda G., Grimaldi D., Kitade O., Krishna K., Klass K.-D., Maekawa K., Miura T., Thompson G.J. Comment. Save Isoptera: a comment on Inward et al. (англ.) // Biology Letters : journal. — Royal Society Publishing, 2007. — Vol. 3, no. 5. — P. 562—563. — doi:10.1098/rsbl.2007.0264. — PMID 17698448. — PMC 2391185.
- ↑ Thomas Bourguignon, Nathan Lo. Termites: Phylogeny and Classification // Encyclopedia of Social Insects (англ.) / Christopher K. Starr (ed.). — Springer Nature Switzerland AG, 2020. — P. 1—6. — ISBN 978-3-319-90306-4. — doi:10.1007/978-3-319-90306-4.
- ↑ Zhao Z. P., Eggleton P., Yin X. C., Gao T. P., Shih C. K. & Ren D. The oldest known mastotermitids (Blattodea: Termitoidae) and phylogeny of basal termites (англ.) // Systematic Entomology : журнал. — John Wiley & Sons, Inc., 2019. — Vol. 44, no. 3. — P. 612—623. — ISSN 0307-6970. — doi:10.1111/syen.12344.
- ↑ Bechly G. 2007. Isoptera: termites. The Crato Fossil Beds of Brazil: Window into an Ancient World 249—262
- ↑ Engel M. S. Two new termites in Baltic amber (Isoptera) (англ.) // Journal of the Kansas Entomological Society : журнал. — 2008. — Vol. 81, no. 3. — P. 194—203. — doi:10.2317/JKES-0802.01.1.
- ↑ 1 2 Engel M. S., Grimaldi, D. A. & Krishna K. Primitive termites from the Early Cretaceous of Asia (Isoptera) (англ.) // Stuttgarter Beitrage zur Naturkunde, Serie B, Geologie und Palaontologie : журнал. — 2007. — Vol. 371. — P. 1—32.
- ↑ Zhao, Z. P., Yin, X. C., Shih, C. K., Gao, T. P. & Ren, D. (2020) Termite colonies from mid-Cretaceous Myanmar demonstrate their early eusocial lifestyle in damp wood. National Science Review, 7 (2), 381—390. https://doi.org/10.1093/nsr/nwz141
- ↑ Engel, M. S., Barden, P., Riccio, M. L. & Grimaldi, D. A. (2016) Morphologically specialized termite castes and advanced sociality in the Early Cretaceous. Current Biology, 26 (4), 522—530. https://doi.org/10.1016/j.cub.2015.12.061
- ↑ Jouault, C., Legendre, F., Grandcolas, P. & Nel, A. (2021) Revising dating estimates and the antiquity of eusociality in termites using the fossilized birth-death process. Systematic Entomology, 46 (3), 592—610. https://doi.org/10.1111/syen.12477
- ↑ Kohler, T; Dietrich, C; Scheffrahn, RH; Brune, A (2012). "High‐resolution analysis of gut environment and bacterial microbiota reveals functional compartmentation of the gut in wood‐feeding higher termites (Nasutitermes spp.)". Applied and Environmental Microbiology. 78 (13): 4691—4701. doi:10.1128/aem.00683-12. PMC 3370480. PMID 22544239.
Литература
- Жужиков Д. П. Термиты СССР. — М.: Издательство Моск. ун-та, 1979. — 225 с.
- Жужиков Д. П. . Особенности структуры и регуляции развития в семье термитов. Чтения памяти Н. А. Холодковского: Докл. на 38 ежегод. чтении, 4 апреля 1985 г. — Л.: Наука, 1986. — С. 74—105.
- Халифман И. А. Отступившие в подземелье. — М.: Детгиз, 1961. — 192 с. — (Школьная библиотека). — 30 000 экз.
- Халифман И. А. Пароль скрещенных антенн. — М.: Детская литература, 1967. — 414 с. — 75 000 экз.
- Халифман И. А. Шмели и термиты. — М.: Детская литература, 1972. — 320 с. — 100 000 экз.
- Эшерих К. Термиты или белые муравьи. — СПб., 1910.
- Abe, T.; D. E. Bignell; M. Higashi, eds. Termites: Evolution, Sociality, Symbiosis, Ecology. — Dordrecht: Kluwer Academic Publishing, 2010. — С. 1—466. — ISBN 978-90-481-5476-0 (Print) 978-94-017-3223-9 (Online).
- Bignell, David Edward; Yves Roisin, Nathan Lo. Biology of Termites: a Modern Synthesis. — Dordrecht: Springer Netherlands, 2010. — ISBN 978-90-481-3977-4..
- Schmid-Hempel P. Parasites in Social Insects. — New Jersey: Princeton University Press, 1998. — ISBN 978-0-691-05924-2.
- Termites: evolution, sociality, symbioses, ecology / Ed. by Takuya Abe (01.03.1945-27.03.2000), David Edward Bignell, Masahiko Higashi (11.08.1954-27.03.2000 ). — Dordrecht, Netherlands: Kluwer Academic Publishers (Springer Netherlands), 2000. — 467 с. — ISBN 978-94-017-3223-9. — doi:10.1007/978-94-017-3223-9_20.
- Donovan, S. E., D. T. Jones, W. A. Sands and P. Eggleton. 2000. The morphological phylogenetics of termites (Isoptera). — Biological Journal of the Linnean Society (The Linnean Society of London.). 70(3):467—513.
- Engel Michael, David A. Grimaldi and Kumar Krishna. 2009. Termites (Isoptera): Their Phylogeny, Classification, and Rise to Ecological Dominance. American Museum Novitates. 3650:1—27. [1]
- Engel, M. and K. Krishna. 2004. Family-group names for termites (Isoptera). — American Museum Novitates 3432:1—9.
- Grassé, P. P. 1986. Termitologia. Comportement, Sociatilé, Ecologie, Evolution, Systematique. — Paris, Masson. — V.3, 715 p.
- Krishna, Kumar.; Grimaldi, David A.; Krishna, Valerie.; Engel, Michael S. Treatise on the Isoptera of the world (англ.) // Bulletin of the American Museum of Natural History[англ.] : Журнал. — Нью-Йорк: American Museum of Natural History, 2013. — Vol. 377 (vol. 1—7). — P. 1—2704. — ISSN 0003-0090.
Ссылки
- Энциклопедия Кругосвет — Термиты
- Termite Laboratory — University of Brasília
- Статья, посвящённая происхождению термитов
- Термиты как одна из экосистем
- Книга «Общественные насекомые» Майкл Брайен
- Беляева Н. В. (2005). Вредная деятельность термитов.
- Война миров: термиты. Журнал «Чайка».
- Войны термитов могут объяснить эволюцию социальных насекомых
- Термитология на www.isoptera.ufv.br (порт.)
| Таксономия | ||||
|---|---|---|---|---|