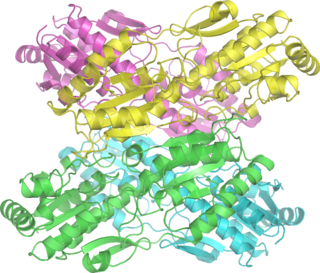
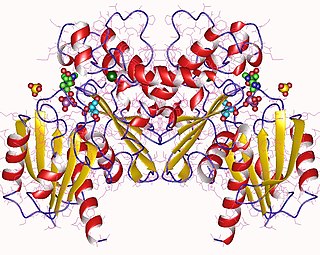
Транскетолазы
 | ксилулозо-5-фосфат | + |  | D-рибозо-5-фосфат |
 | D-глицеральдегид-3-фосфат | + |  | седогептулозо-7-фосфат |
Транскетолазы — группа ферментов пентозофосфатного пути и цикла Кальвина. Они катализирует две важные реакции, которые действуют в противоположных направлениях в этих двух путях.
Транскетолазы переносят двухуглеродную группу, включающую 1-й и 2-й атомы углерода кетозы, на альдегидный углерод альдозного сахара. Происходит превращение кетосахара в альдозу, содержащую на два атома углерода меньше, и одновременное превращение альдосахара в кетозу, содержащую на два атома углерода больше.
Транскетолазы катализируют перенос двухуглеродной группы с ксилулозо-5-фосфата на рибозо-5-фосфат с образованием семиуглеродной кетозы седогептулозо-7-фосфата и альдозы глицеральдегид-3-фосфата. В другой реакции ксилулозо-5-фосфат служит донором «активного гликоальдегида». Роль акцептора выполняет эритрозо-4-фосфат. Продуктами этой реакции являются фруктозо-6-фосфат и глицеральдегид-3-фосфат.
У млекопитающих транскетолазы соединяют пентозофосфатный путь с гликолизом, переводя избыток сахарофосфатов в основной метаболический путь углеводов. Его присутствие необходимо для производства НАДФН, особенно в тканях активно участвует в процессах биосинтеза, таких как синтез жирных кислот в печени и молочных железах, и для стероидный синтез в печени и надпочечниках. Главными кофакторами этого процесса являются тиаминдифосфат и кальций.
Транскетолазы обильно экспрессируются у млекопитающих в роговице стромальными кератоцитами и клетками эпителия. Считается, что они являются одним из роговичных кристаллинов[1].
Распространение
Транскетолазы присутствуют у в большого количества организмов, включая бактерий, растений и млекопитающих. У человека также есть гены, кодирующие белки с транскетолазной активностью:
- ТКТ (транскетолаза)
- TKTL1 (транскетолазо-подобный белок 1)
- TKTL2 (транскетолазо-подобный белок 2)
Структура
Вход в активный центр этого фермента состоит в основном из боковых групп аргинина, гистидина, серина и аспарагиновой кислоты, а глутамат играет второстепенную роль. Эти боковые цепи, а именно Arg359, Arg528, His469 и Ser386, сохраняются внутри каждого фермента транскетолазы и взаимодействуют с фосфатной группой донорных и акцепторных субстратов. Поскольку субстратный канал настолько узкий, донорные и акцепторные субстраты не могут связываться одновременно. Кроме того, субстраты при связывании в активном центре изменяют свою конформацию на более вытянутую, чтобы уместиться этот узкий канал.
Хотя транскетолазы способны связывать различные субстраты, например фосфорилированные и не фосфорилированные моносахариды в том числе кетозы и фруктоза, рибоза и так далее, они имеет высокую стереоспецифичность по отношению к кетозам с транс-положением гидроксильных групп при атомах С-3 и С-4[2]. Также они стабилизируют субстрат в активном центре при помощи остатков Asp477, His30, и His263. Нарушение этой конфигурации, размещение гидроксильных групп или их стереохимии, приводит к нарушению водородных связей между аминокислотными остатками и субстратом, что приводит к более низкому сродству к субстрату.
В первой половине этого пути, His263 используется для эффективного отщепления С3 гидроксильных протонов, что позволяет 2-углеродному фрагменту отщепляется от фруктозы 6-фосфата[3]. Кофактор, необходимый для этого шага — тиаминпирофосфат. Связывание тиамина с ферментом не приводит к каким бы то ни было серьёзным конформационным изменениям фермента; наоборот, фермент состоит из двух гибких петель в активном центре, которые делают тиаминпирофосфат доступными для связывания[2].
Механизм
Каталитический механизм начинается с депротонирования тиазолового кольца тиаминприофосфата. Образовавшийся карбанион связывается с карбонильной группой донорного субстрата таким образом, что расщепляется связь между C-2 и C-3 атомами. Этот двухуглеродный фрагмент остается ковалентно связан С-2 углеродом тиаминпирофосфата. После этого донорный субстрат высвобождается, и акцептором субстрат входит в активный сайт, где фрагмент, связанный с α-β-дигидроксиэтилтиаминпирофосфат переносится на акцептор[2].

![]()
Были проведены эксперименты, проверявшие эффект замены аланина на аминокислоты у входа в активный центр, Arg359, Arg528, и His469, которые взаимодействуют с фосфатной группой субстрата. Эта замена приводит к образованию фермента с нарушенной каталитической активностью[2].
Роль в заболеваниях
Транскетолазная активность снижается при дефиците тиамина, в основном по причине недоедания. С дефицитом тиамина связан ряд заболеваний, в том числе и бери-бери, биотин-тиамин-отзывчивая болезнь базальных ганглиев[4], синдром Вернике-Корсакова и другие.
Не было обнаружено каких-либо специфических мутаций связанных с синдромом Вернике-Корсакова[5], но есть указание, что дефицит тиамина приводит к развитию этого синдрома только у тех, чьи транскетолазы имеют пониженное сродство к тиамину[6]. таким образом, активность транскетолазы сильно затруднена, и, как следствие, тормозится весь пентозофосфатный путь[7].
Диагностика
Транскетолазная активность эритроцитов уменьшается при дефиците тиамина (витамина В1), что используется для диагностики энцефалопатии Вернике и других синдромов, связанных с недостатком витамина B1, если есть сомнения в диагнозе[8]. Помимо базовой ферментативной активностью (которая может быть нормальной даже при дефиците), усиление активности фермента после добавления тиаминпирофосфата может быть использовано для диагностики дефицита тиамина (0-15 % нормальных, 15-25 % дефицит, >25 % тяжелая недостаточность)[9].
Список литературы
- ↑ Sax C.M., Kays W.T., Salamon C., Chervenak M.M., Xu Y.S., Piatigorsky J. Transketolase gene expression in the cornea is influenced by environmental factors and developmentally controlled events (англ.) // Cornea : journal. — 2000. — November (vol. 19, no. 6). — P. 833—841. — doi:10.1097/00003226-200011000-00014. — PMID 11095059.
- ↑ 1 2 3 4 Nilsson U., Meshalkina L., Lindqvist Y., Schneider G. Examination of substrate binding in thiamin diphosphate-dependent transketolase by protein crystallography and site-directed mutagenesis (англ.) // J. Biol. Chem. : journal. — 1997. — January (vol. 272, no. 3). — P. 1864—1869. — doi:10.1074/jbc.272.3.1864. — PMID 8999873.
- ↑ Wikner C., Nilsson U., Meshalkina L., Udekwu C., Lindqvist Y., Schneider G. Identification of catalytically important residues in yeast transketolase (англ.) // Biochemistry : journal. — 1997. — December (vol. 36, no. 50). — P. 15643—15649. — doi:10.1021/bi971606b. — PMID 9398292.
- ↑ Biotin-Thiamine-Responsive Basal Ganglia Disease - GeneReviews® - NCBI Bookshelf. Дата обращения: 3 октября 2017. Архивировано 9 мая 2018 года.
- ↑ McCool B.A., Plonk S.G., Martin P.R., Singleton C.K. Cloning of human transketolase cDNAs and comparison of the nucleotide sequence of the coding region in Wernicke-Korsakoff and non-Wernicke-Korsakoff individuals (англ.) // J. Biol. Chem. : journal. — 1993. — January (vol. 268, no. 2). — P. 1397—1404. — PMID 8419340.
- ↑ Blass J.P., Gibson G.E. Abnormality of a thiamine-requiring enzyme in patients with Wernicke-Korsakoff syndrome (англ.) // N. Engl. J. Med. : journal. — 1977. — Vol. 297, no. 25. — P. 1367—1370. — doi:10.1056/NEJM197712222972503. — PMID 927453.
- ↑ Cox, Michael; Nelson, David R.; Lehninger, Albert L. Lehninger principles of biochemistry. — San Francisco: W.H. Freeman[англ.], 2005. — ISBN 0-7167-4339-6.
- ↑ Smeets E.H., Muller H., de Wael J. A NADH-dependent transketolase assay in erythrocyte hemolysates (англ.) // Clin. Chim. Acta[англ.] : journal. — 1971. — July (vol. 33, no. 2). — P. 379—386. — doi:10.1016/0009-8981(71)90496-7. — PMID 4330339.
- ↑ Doolman R., Dinbar A., Sela B.A. Improved measurement of transketolase activity in the assessment of "TPP effect" (англ.) // Eur J Clin Chem Clin Biochem : journal. — 1995. — July (vol. 33, no. 7). — P. 445—446. — PMID 7548453.