Ферме́нты, или энзи́мы , — обычно сложные белковые соединения, РНК (рибозимы) или их комплексы, ускоряющие химические реакции в живых системах. Каждый фермент, свернутый в определённую структуру, ускоряет соответствующую химическую реакцию: реагенты в такой реакции называются субстратами, а получающиеся вещества — продуктами. Ферменты специфичны к субстратам: АТФ-аза катализирует расщепление только АТФ, а киназа фосфорилазы фосфорилирует только фосфорилазу.

Ци́кл трикарбо́новых кисло́т (сокр. ЦТК, цикл Кре́бса, цитра́тный цикл, цикл лимо́нной кислоты́) — центральная часть общего пути катаболизма, циклический биохимический процесс, в ходе которого ацетильные остатки (СН3СО-) окисляются до диоксида углерода (CO2). При этом за один цикл образуется 2 молекулы CO2, 3 НАДН, 1 ФАДH2 и 1 ГТФ (или АТФ). Электроны, находящиеся на НАДН и ФАДH2, в дальнейшем переносятся на дыхательную цепь, где в ходе реакций окислительного фосфорилирования образуется АТФ.

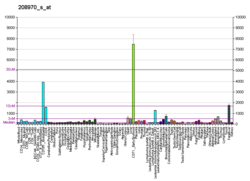
Глюкокиназа — это IV изотип фермента гексокиназы, в основном присутствующий в гепатоцитах, а также в клетках поджелудочной железы. Катализирует фосфорилирование шестиатомных сахаров (гексоз), в частности D-глюкозы, D-маннозы, D-фруктозы, D-глюкозамина, сорбитола.

Транспортная РНК, тРНК — рибонуклеиновая кислота, обеспечивающая взаимодействие аминокислоты, рибосомы и матричной РНК (мРНК) в ходе трансляции. Имеет типичную длину от 73 до 93 нуклеотидов и размеры около 5 нм. тРНК, будучи ковалентно связаны с остатком аминокислоты, принимает непосредственное участие в наращивании полипептидной цепи, специфически присоединяясь к кодону мРНК и обеспечивая необходимую для образования новой пептидной связи конформацию комплекса.
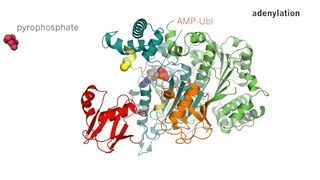
Ферментативный катализ — это увеличение скорости процесса с помощью биологической молекулы, «фермента». Большинство ферментов представляют собой белки, и большинство таких процессов представляют собой химические реакции. Внутри фермента катализ обычно происходит в локализованном центре, называемом активным центром.

РНК-полимераза II — фермент эукариот, который катализирует транскрипцию ДНК, синтезирует предшественников мРНК и большинство мяРНК и микроРНК. Эта полимераза представляет собой комплекс массой 550 кДа, состоящий из 12 субъединиц. РНК-полимераза II является наиболее изученным типом РНК-полимеразы. Ей необходим широкий спектр транскрипционных факторов для того, чтобы связываться с генами выше промоторов и начинать транскрипцию.
Ацил-КоА-дегидрогеназы, также ACADs — семейство ферментов из класса оксидоредуктаз, которые катализируют реакции переноса протона (дегидрогенизация) от субстрата — ацил-КоА жирной кислоты на электрон-переносящий флавопротеин (FAD), участвуют в процессе β-окисления. Результатом реакции является образование двойной связи расположенной между атомами С2 (α) и С3 (β) в молекуле тиоэфира субстрата (ацил-КоА).

Серин-С-пальмитоилтрансфераза, также серин-пальмитоилтрансфераза, 3-оксосфинганин синтаза (англ. serine C-palmitoyltransferase, сокр. SPT) — фермент (КФ 2.3.1.50), из семейства ацилтрансфераз (класс трансферазы), катализирующий реакцию переноса ацильной группы на молекулу субстрата — серина, по уравнению:
- пальмитоил-CoA + серин
 CoA-SH + 3-дегидро-D-сфинганин + CO2.
CoA-SH + 3-дегидро-D-сфинганин + CO2.

НАДФ-зависимая декарбоксилирующая малатдегидрогеназа или НАДФ-малик-энзим (НАДФ-МЭ) представляет собой фермент, катализирующий химическую реакцию в присутствии двухвалентных ионов металлов:
- (S)-малат + НАДФ+ -> пируват + CO2 + НАДФН

B-Raf, или c-RAF — цитозольная серин/треониновая протеинкиназа семейства MAP3K. Продукт протоонкогена B-Raf BRAF.
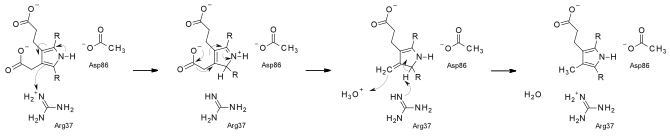
Ферментный промискуитет (неразборчивость) — это способность фермента катализировать случайную побочную реакцию в дополнение к своей основной реакции. Хотя ферменты являются чрезвычайно специфическими катализаторами, они часто могут выполнять побочные реакции в дополнение к своей основной природной каталитической активности. Побочная активность фермента обычно протекает медленнее по сравнению с основной деятельностью и находится под нейтральным отбором. Несмотря на то, что обычно эти активности физиологически нерелевантны, в условиях нового избирательного давления эти виды деятельности могут принести пользу, тем самым побуждая эволюцию ранее побочных активностей стать новым основным видом деятельности. Примером этого является хлоргидролаза атразина (кодируется atzA) Pseudomonas sр., произошедшая из меламиндезаминазы, которая имеет очень небольшую побочную активность в отношении атразина, химического вещества, созданного руками человека.
Дегидрогеназный комплекс разветвлённых α-кетокислот, также дегидрогеназа α-кетокислот с разветвлённой цепью — мультиферментный комплекс, локализованный на внутренней мембране митохондрий. Данный комплекс катализирует реакцию окислительного декарбоксилирования разветвлённых α-кетокислот с короткой углеродной цепью. Дегидрогеназный комплекс структурно аналогичен пируватдегидрогеназному и α-кетоглутаратдегидрогеназному комплексам, образуя вместе с ними семейство митохондриальных ферментов, участвующих в окислительном декарбоксилировании α-кетокислот.

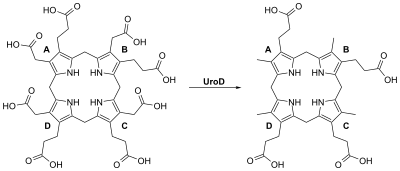
Уропорфириноген III синтаза — фермент из класса лиазы, который катализирует реакцию циклизации линейных молекул гидроксиметилбилана в уропорфириноген III. При катализе происходит инверсия конечного пиррольного звена линейной молекулы гидроксиметилбилана, связывая его с первым пиррольным звеном, тем самым образуя крупную макроциклическую структуру, уропорфириноген III.

Биосинтез гема — метаболический многоступенчатый процесс образования молекул гема из его предшественников. Гем представляет собой плоский железосодержащий макроциклический комплекс, основу которого составляет четыре пиррольных кольца (тетрапиррол), объединённые между собой метениловыми мостиками (=СH-) и координированные двухвалентным железом (Fe2+). Гем имеет несколько типов, они отличаются как структурно, так и функционально. Но наиболее распространённым типом гема, является — гем Б.

Протопорфириноген IX оксидаза, также протопорфириноген оксидаза (сокр. ППГ-IX-оксидаза) — фермент (1.3.3.4) из семейства оксидазы (класс оксидоредуктазы), катализирует реакцию окисления протопорфориногена IX до протопорфирина IX, путём отщепления 6 атомов водорода по схеме:
- протопорфириноген IX + 3O2 → протопорфирин IX + 3H2O2

Альдолаза B, также фруктозо-бисфосфат альдолаза B или реже печёночный тип альдолазы — фермент из семейства фруктозо-1,6-бисфосфат альдолазы I класса, является одним из трёх изоферментов. Альдолаза B играет ключевую роль в углеводном обмене: в процессах гликолиза и глюконеогенеза, а также в метаболизме фруктозы. Фермент катализирует реакцию альдольного (негидролитического) расщепления связей C-C в молекуле фруктозо-1,6-бисфосфата (Ф-1,6-БФ) до дигидроксиацетонфосфата (ДАФ) и глицеральдегид-3-фосфата (ГАФ), обратную реакцию — альдольную конденсацию, а также расщепление фруктозо-1-фосфата на глицеральдегид и дигидроксиацетонфосфат, и их альдольную конденсацию во фруктозо-1-фосфат. Схема реакции, катализируемой альдолазой B:
- фруктозо-1,6-бисфосфат
 дигидроксиацетонфосфат + глицеральдегид-3-фосфат
дигидроксиацетонфосфат + глицеральдегид-3-фосфат - фруктозо-1-фосфат дигидроксиацетонфосфат + глицеральдегид.

Альдолаза A, также фруктозо-бисфосфат альдолаза A или реже мышечный тип альдолазы — фермент из семейства фруктозо-1,6-бисфосфат альдолазы I класса, является одним из трёх изоферментов. Альдолаза А играет ключевую роль в углеводном обмене: в глюконеогенезе и гликолизе в скелетных мышцах и эритроцитах. Фермент катализирует обратимую реакцию альдольного расщепления (негидролитического) и конденсации молекулы фруктозо-1,6-бисфосфата по реакции:
- фруктозо-1,6-бисфосфат дигидроксиацетонфосфат + глицеральдегид-3-фосфат.
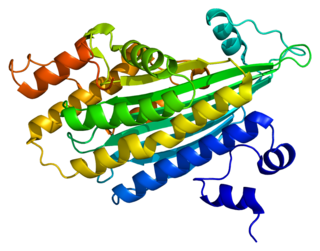
Копропорфириноген III оксидаза, сокр. КПГ-III-оксидаза (англ. Coproporphyrinogen-III oxidase, сокр. CPOX) — фермент (КФ 1.3.3.3) семейства оксидаз (класс оксидоредуктазы), катализирующий реакцию окислительного декарбоксилирования копропорфориногена III в протопорфириноген IX. Схема реакции:
- копропорфириноген III + 2O2 + 2H+ → протопорфириноген IX + 2H2O + 2CO2

Аргининосукцинат-синтетаза — фермент, из семейства N-C лигаз, катализирующий реакцию синтеза аргининосукцината путём присоединения (конденсации) молекул аспартата и цитруллина, с использованием энергии гидролиза макроэргических связей молекул ATP. Схема реакции: