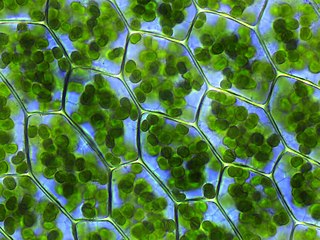
Устьице

У́стьице (лат. stoma, от греч. στόμα — «рот, уста») в ботанике — высокоспециализированное образование эпидермиса растений, состоящее из двух замыкающих клеток и устьичной щели (апертуры) между ними, через которую осуществляется газообмен и транспирация.
История исследований
Хотя учёные давно знали об испарении воды поверхностью листа, первым, кто наблюдал устьица, был итальянский натуралист Марчелло Мальпиги, который это открытие опубликовал в 1675 году в своей работе Anatomia plantarum. Однако он не понял их настоящую функцию. В то же время его современник Неемия Грю развил гипотезу об участии устьиц в вентиляции внутренней среды растения и сравнил их с трахеями насекомых. Прогресс в изучении наступил в XIX веке, и тогда же, в 1827 году, швейцарским ботаником Декандолем было впервые использовано слово «stoma». Изучением устьиц в то время занимались Гуго фон Моль, который открыл основной принцип открывания устьиц, и Симон Швенденер, классифицировавший устьица по типу их строения.
Некоторые аспекты функционирования устьиц продолжают интенсивно изучаться и в настоящее время; материалом в основном служат Коммелина обыкновенная (Commelina communis), Боб садовый (Vicia faba), Кукуруза сахарная (Zea mays)[1].
Строение

Размеры устьица (длина) колеблются в пределах 0,01—0,06 мм (крупнее бывают устьица полиплоидных растений и у листьев, растущих в тени). Самые крупные устьица были обнаружены у вымершего растения Zosterophyllum, 0,12 мм (120 мкм)[1]. Устьице состоит из пары специализированных клеток, называемых замыкающими (cellulae claudentes), которые регулируют степень открытости устьица, между ними располагается устьичная щель (porus stomatalis). Стенки замыкающих клеток утолщены неравномерно: направленные к щели (брюшные) толще стенок, направленных от щели (спинных). Щель может расширяться и сужаться, регулируя транспирацию и газообмен. Когда воды мало, замыкающие клетки плотно прилегают друг к другу и устьичная щель закрыта. Когда воды в замыкающих клетках много, то она давит на стенки и более тонкие стенки растягиваются сильнее, а более толстые втягиваются внутрь, между замыкающими клетками появляется щель[2]. Под щелью расположена подустьичная (воздушная) полость, через которую непосредственно и происходит газообмен. Воздух, содержащий диоксид углерода (углекислый газ) и кислород, проникает внутрь ткани листа через устьица, и далее используется в процессе фотосинтеза и дыхания. Избыточный кислород, произведённый в процессе фотосинтеза выходит обратно в окружающую среду через устьица. Также в процессе транспирации через устьица выделяются пары воды. Клетки эпидермиса, примыкающие к замыкающим, получили название сопровождающих (побочных, соседних, околоустьичных). Они участвуют в движении замыкающих клеток. Замыкающие и сопровождающие клетки образуют устьичный комплекс (устьичный аппарат). Наличие или отсутствие устьиц (видимые части устьиц называют устьичными линиями) часто используют при классификации растений.
Типы устьиц
Число сопровождающих клеток и их расположение относительно устьичной щели позволяют выделить ряд типов устьиц:
- аномоцитный — сопровождающие клетки не отличаются от остальных клеток эпидермиса, тип весьма обычен для всех групп высших растений, за исключением хвойных;
- диацитный — характеризуется только двумя сопровождающими клетками, общая стенка которых находится под прямым углом к замыкающим клеткам;
- парацитный — сопровождающие клетки располагаются параллельно замыкающим и устьичной щели;
- анизоцитный — замыкающие клетки окружены тремя неравными сопровождающими, одна из которых заметно крупнее или мельче остальных, такой тип обнаружен только у цветковых растений;
- тетрацитный — четыре сопровождающие клетки, характерен для однодольных;
- энциклоцитный — сопровождающие клетки образуют узкое кольцо вокруг замыкающих клеток;
- актиноцитный — несколько сопровождающих клеток радиально расположены вокруг устьичных клеток, напоминая лучи звезды;
- перицитный — замыкающие клетки окружены одной побочной сопровождающей клеткой, устьице не соединено с сопровождающей клеткой антиклинальной клеточной стенкой;
- десмоцитный — замыкающие клетки окружены одной сопровождающей клеткой, устьице соединено с ней антиклинальной клеточной стенкой;
- полоцитный — замыкающие клетки окружены одной сопровождающей не полностью: к одному из устьичных полюсов примыкает одна или две эпидермальные клетки; устьице прикреплено к дистальной стороне единственной сопровождающей клетки, имеющей U-образную или подковообразную форму;
- стефаноцитный — устьице, окружённое четырьмя или более (обычно пять-семь) слабодифференцированными сопровождающими клетками, образующими более или менее отчётливую розетку;
- латероцитный — такой тип устьичного аппарата рассматривается большинством ботаников как простая модификация аномоцитного типа.
У двудольных распространённым является парацитный тип устьиц. Замыкающие клетки почковидной (бобовидной) формы — такими они видны с поверхности листа — несут хлоропласты, тонкие неутолщённые участки оболочки образуют выступы (носики), закрывающие устьичную щель.
Наружные стенки замыкающих клеток обычно имеют выросты, что хорошо видно на поперечном разрезе устьица. Пространство, ограниченное этими выростами, называют передним двориком. Нередко аналогичные выросты наблюдаются и у внутренних оболочек замыкающих клеток. Они образуют задний дворик, или внутренний, соединённый с крупным межклетником — подустьичной полостью.
Из однодольных парацитное строение устьиц отмечено у злаков. Замыкающие клетки имеют гантелевидную форму — сужены в средней части и расширены на обоих концах, при этом стенки расширенных участков очень тонкие, а в средней части замыкающих клеток сильно утолщены. Хлоропласты располагаются в пузыревидных окончаниях клеток.
Для одних видов растений характерен только один тип устьичного аппарата, для других — два и несколько даже в пределах одной листовой пластинки[3].
Движение замыкающих клеток
Механизм движения замыкающих клеток весьма сложен и неодинаков у разных видов. У большинства растений при неодинаковом водоснабжении в ночные часы, а иногда и днём тургор в замыкающих клетках понижается, и устьичная щель замыкается, снижая тем самым уровень транспирации. С повышением тургора устьица открываются. Считают, что главная роль в изменении тургора принадлежит ионам калия. Существенное значение в регуляции тургора имеет присутствие в замыкающих клетках хлоропластов. Первичный крахмал хлоропластов, превращаясь в сахар, повышает концентрацию клеточного сока. Это способствует притоку воды из соседних клеток и повышению тургорного давления в замыкающих клетках[4].
Расположение устьиц
Двудольные растения, как правило, в нижней части листа имеют больше устьиц, чем в верхней. Это объясняется тем, что верхняя часть горизонтально-расположенного листа, как правило, лучше освещена, и меньшее количество устьиц в ней препятствует избыточному испарению воды. Листья с устьицами, расположенными на нижней стороне, называются гипостоматическими.
У однодольных растений наличие устьиц в верхней и нижней части листа различно. Очень часто листья однодольных растений расположены вертикально, и в этом случае количество устьиц на обеих частях листа может быть одинаково. Такие листья называются амфистоматическими.
У плавающих листьев на нижней части листа устьица отсутствуют, так как они могут впитывать воду через кутикулу. Листья с устьицами, расположенными на верхней стороне, называются эпистоматическими. У подводных листьев устьица отсутствуют совсем.
Устьица хвойных растений обычно спрятаны глубоко под эндодермой, что позволяет сильно снизить расход воды на испарение, как зимой, так и летом во время засухи.
У мхов (исключение — антоцеротовые) настоящие устьица отсутствуют.
Устьица также различаются по уровню расположения относительно поверхности эпидермиса. Некоторые из них расположены вровень с другими эпидермальными клетками, другие подняты выше или погружены ниже поверхности. У однодольных, листья которых растут преимущественно в длину, устьица образуют правильные параллельные ряды, тогда как у двудольных они располагаются беспорядочно.
Углекислый газ
Так как углекислый газ является одним из ключевых реагентов в процессе фотосинтеза, у большинства растений устьица в дневное время открыты. Проблема состоит в том, что при входе воздух смешивается с парами воды, испаряющимися из листа, и поэтому растение не может получить углекислый газ, одновременно не потеряв некоторое количество воды. У многих растений существует защита от испарения воды в виде закупоривающих устьица восковых отложений.
Примечания
- ↑ 1 2 WILLMER, Colin Michael; FRICKER, Mark. Stomata Архивная копия от 1 июля 2020 на Wayback Machine. [s.l.] : Springer, 1996.
- ↑ Тема 3. Дата обращения: 6 декабря 2012. Архивировано из оригинала 16 января 2012 года.
- ↑ Лотова Л. И., Тимонин А. К. Сравнительная анатомия высших растений. — М.: Изд-во МГУ, 1989. — 80 с.
- ↑ В. Р. Пикеринг. Устьица // Биология. Школьный курс в 120 таблицах. — М.: «АСТ-ПРЕСС», 1997. — С. 44. — 25 000 экз. — ISBN 5-7805-0179-3.
Литература
- Устьица растений // Энциклопедический словарь Брокгауза и Ефрона : в 86 т. (82 т. и 4 доп.). — СПб., 1890—1907.
- Атлас по анатомии растений: учеб. пособие для вузов / Бавтуто Г. А., Ерёмин В. М., Жигар М. П.. — Мн.: Ураджай, 2001. — 146 с. — (Учеб. и учеб. пособия для вузов). — ISBN 985-04-0317-9.
- Colin Michael Willmer, Mark Fricker. Stomata. — Chapman & Hall, 1995. — ISBN 0412574306.
| В библиографических каталогах |
|---|