
Клеточный рецептор — молекула на поверхности клетки, клеточных органелл или растворенная в цитоплазме. Специфично реагирует изменением своей пространственной конфигурации на присоединение к ней молекулы определённого химического вещества, передающего внешний регуляторный сигнал и, в свою очередь, передает этот сигнал внутрь клетки или клеточной органеллы, нередко при помощи так называемых вторичных посредников или трансмембранных ионных токов.

В биологии активный центр — это область фермента, где молекулы субстрата связываются и подвергаются химической реакции. Активный центр состоит из аминокислотных остатков, которые образуют временные связи с субстратом, и остатков, которые катализируют реакцию этого субстрата. Хотя активный центр занимает только ~ 10-20 % от объёма фермента он является наиболее важной частью, поскольку он непосредственно катализирует химическую реакцию. Обычно активный центр состоит из трех-четырех аминокислот, в то время как другие аминокислоты в белке необходимы для поддержания его третичной структуры.

Антагонист в биохимии и фармакологии — подтип лигандов к клеточным рецепторам. Лиганд, обладающий свойствами антагониста рецепторов — это такой лиганд, который блокирует, снижает или предотвращает вызываемые связыванием агониста с рецептором физиологические эффекты. Сам же он при этом не обязан производить какие-либо физиологические эффекты вследствие своего связывания с рецептором. Таким образом, антагонисты рецепторов имеют сродство (аффинность) к данному конкретному типу рецепторов, но, исходя из строгого определения, не имеют собственной внутренней агонистической активности по отношению к этому рецептору, и их связывание лишь нарушает взаимодействие [конкурентных] полных или частичных агонистов с рецептором и предотвращает или ингибирует их функцию и их физиологические эффекты. В равной степени антагонисты рецепторов предотвращают и воздействие на рецептор обратных агонистов. Антагонисты рецепторов опосредуют свои эффекты благодаря связыванию либо с активным сайтом рецептора, тем же самым, с которым связывается и физиологический эндогенный агонист, либо с аллостерическими сайтами, или же они могут взаимодействовать с рецептором в уникальных участках связывания, которые не являются нормальными участками связывания эндогенных веществ для данного рецептора и не принимают в норме участия в физиологической регуляции активности данного рецептора.

Рецептор липопротеинов низкой плотности — белок, опосредующий эндоцитоз липопротеинов низкой плотности, обогащённых холестерином. ЛПНП-рецептор представляет собой мембранный белок, специфически распознающий апоВ-100 и апоЕ. За открытие этого важнейшего рецептора липидного метаболизма Майкл Браун и Джозеф Голдштейн получили Нобелевскую премию по физиологии и медицине в 1985 году. Это стало результатом их работ по наследственной гиперхолестеринемии.
Вторичные посредники, или «вторичные мессенджеры» — это внутриклеточные сигнальные молекулы, высвобождаемые в тех или иных внутриклеточных сигнальных каскадах в ответ на стимуляцию тех или иных рецепторов и вызванную ею активацию первичных эффекторных белков. Вторичные посредники, в свою очередь, приводят к активации вторичных эффекторных белков. Это, в свою очередь, запускает каскад тех или иных физиологических изменений, которые могут быть важны для обеспечения таких важных физиологических процессов, как рост, развитие и дифференцировка клеток, активация деления клетки, транскрипция или, наоборот, угнетение транскрипции тех или иных генов, биосинтез тех или иных белков, выделение ею гормонов, нейромедиаторов или цитокинов соответственно типу клетки, изменение биоэлектрической активности клетки, миграция клеток, обеспечение их выживаемости или, наоборот, индукция апоптоза. Вторичные посредники являются инициирующими элементами во множестве внутриклеточных сигнальных каскадов. Вследствие всего этого вторичные посредники играют очень важную роль в жизни клетки, а грубое нарушение работы любой из систем вторичных посредников оказывает неблагоприятное воздействие на клетку.

Факторы транскрипции — белки́, контролирующие процесс синтеза мРНК, а также других видов РНК на матрице ДНК (транскрипцию) путём связывания со специфичными участками ДНК. Транскрипционные факторы выполняют свою функцию либо самостоятельно, либо в комплексе с другими белками. Они обеспечивают снижение (репрессоры) или повышение (активаторы) константы связывания РНК-полимеразы с регуляторными последовательностями регулируемого гена.
Протеинкина́зы — подкласс ферментов киназ (фосфотрансфераз). Протеинкиназы модифицируют другие белки путём фосфорилирования остатков аминокислот, имеющих гидроксильные группы или гетероциклической аминогруппы гистидина.

АМРА-рецептор — ионотропный рецептор глутамата, который передаёт быстрые возбуждающие сигналы в синапсах нервной системы позвоночных. Данные рецепторы также активируются синтетическим аналогом глутамата — аминокислотой АМРА, откуда и получили своё название. АМРА-рецепторы обнаружены практически во всех структурах головного мозга, их считают наиболее распространённым типом рецепторов в нервной системе. Эти рецепторы представляют собой тетрамерные ионные каналы, которые могут состоять из субъединиц четырёх типов. АМРА-рецепторы имеют отношение к развитию некоторых заболеваний центральной нервной системы человека, таких как синдром Мартина — Белл, поэтому их изучению уделяется большое внимание.

В биохимии и фармакологии лиганд — это химическое соединение, которое образует комплекс с той или иной биомолекулой и производит, вследствие такого связывания, те или иные биохимические, физиологические или фармакологические эффекты. В случае связывания лиганда с белком лиганд обычно является малой сигнальной молекулой, связывающейся со специфическим участком связывания на белке-мишени. В случае связывания лиганда с ДНК лиганд обычно также является малой молекулой или ионом, или белком который связывается с двойной спиралью ДНК.

ГАМКА-рецептор — лиганд-зависимый ионный канал в химических синапсах нервной системы, который тормозит передачу нервного возбуждения и управляется с помощью ГАМК. Это, наряду с ГАМКС-рецептором, один из двух ионотропных ГАМК-рецепторов, отвечающий за реакцию организма на гамма-аминомасляную кислоту. Помимо места, связывающего ГАМК, рецепторный комплекс содержит аллостерические сегменты, способные связывать бензодиазепины, барбитураты, этанол, фуросемид, нейростероиды и пикротоксин.
Трансмембранные рецепторы — мембранные белки, которые размещаются, и работают не только во внешней клеточной мембране, но и в мембранах компартментов и органелл клетки. Связывание с сигнальной молекулой происходит с одной стороны от мембраны, а клеточный ответ формируется на другой стороне от мембраны. Таким образом, они играют уникальную и важную роль в межклеточных связях и передаче сигнала.
Сродство́ — способность одного объекта (тела) связываться с другим объектом и образовывать таким образом новый комплексный объект. Понятие сродство применяется как к физическим объектам, так и к сложным молекулам, в том числе и к белкам. Коэффициентом, характеризующим силу сродства, является энергия связи, выделяющаяся или затрачиваемая при возникновении связи объектов, измеряемая в килоджоулях (кДж) или электронвольтах (эВ).

Рибопереключа́тель — элемент, как правило, 5'-нетранслируемой области (5'-НТО) мРНК. Он осуществляет цис-регуляцию мРНК, на которой находится, путём связывания с лигандами — разнообразными малыми молекулами, например, кобаламидом, тиаминпирофосфатом, лизином, глицином, флавинмононуклеотидом, гуанином, аденином и другими. Типичный рибопереключатель включает два основных домена: аптамерный домен, осуществляющий распознавание лиганда и связывание с ним, и платформу экспрессии, которая взаимодействует с белками транскрипции или трансляции. Аптамерный домен и платформа экспрессии перекрываются в области так называемой переключающей последовательности, которая и отвечает за сворачивание РНК в две взаимноисключающие вторичные структуры, за счёт чего и осуществляется регуляция.
Тормозный постсинапти́ческий потенциа́л — это разновидность постсинаптического потенциала, которая приводит к тому, что активность постсинаптического нейрона понижается, и менее вероятным становится возникновение потенциала действия.
Конкурентные антагонисты — подтип лигандов-антагонистов рецепторов, которые обратимо связываются с рецепторами в том же самом участке связывания, что и физиологический эндогенный лиганд-агонист этого рецептора, но не вызывают активации рецептора. Физиологические агонисты и конкурентные антагонисты в этом случае «конкурируют» за связывание с одним и тем же участком связывания рецепторов. После того, как конкурентный антагонист свяжется с участком связывания рецептора, он предотвращает связывание с ним же агониста. Однако конкурентный антагонист не может ни «вытеснить» уже связавшийся с рецептором агонист из связи, ни предотвратить воздействие уже связавшегося агониста на клетку. Конечный результат конкуренции агонистов и антагонистов — и таким образом конечный уровень активности рецепторной системы — определяется соотношением молярных концентраций, относительным сродством к рецепторам и соотношением относительной внутренней агонистической активности агонистов и антагонистов. Поскольку высокие концентрации конкурентного антагониста повышают процент занятости рецепторов этим антагонистом, для достижения того же самого процента занятости рецепторов агонистом в этих условиях — и получения того же самого физиологического ответа — потребуются более высокие концентрации агониста, и наоборот — при более высоких концентрациях агониста требуется больше конкурентного антагониста для функциональной «блокады» рецепторов. В функциональных исследованиях конкурентные антагонисты вызывают параллельный сдвиг кривой «доза агониста-эффект» вправо, без изменения максимальной величины физиологического ответа.
Необратимые антагонисты ковалентно связываются с рецептором-мишенью, необратимо меняя его пространственную конфигурацию и тем самым необратимо инактивируют его. Необратимые антагонисты, как правило, не могут быть ферментативно удалены из связи с рецептором. Таким образом, продолжительность физиологического воздействия необратимого антагониста определяется не традиционной кинетикой связывания рецептора с лигандом, а скоростью оборота рецепторов — скоростью процесса физиологического «выведения из строя» и удаления с поверхности мембраны клетки старых, «деградировавших» от времени рецепторов и скоростью биосинтеза клеткой и выведения на поверхность клеточной мембраны новых рецепторов взамен старых, деградировавших. Примером необратимого антагониста α-адренорецепторов является феноксибензамин, который ковалентно и необратимо связывается с α-адренорецепторами, предотвращая тем самым связывание с ними адреналина и норадреналина. Инактивация рецепторов необратимым агонистом обычно приводит к снижению или уменьшению максимально возможного физиологического ответа на максимальную агонистическую стимуляцию. Помимо этого, в системах, в которых есть рецепторный резерв, также может наблюдаться сдвиг кривой «доза-эффект» вправо, подобный сдвигу кривой вправо, наблюдаемому при воздействии конкурентных антагонистов. Отмывание культуры клеток, которые подверглись воздействию антагониста, от остатков антагониста, обычно позволяет различить воздействие неконкурентного антагониста от воздействия необратимого антагониста, поскольку действие неконкурентных антагонистов кратковременно и обратимо, и после отмывания клеток от антагониста эффективность воздействия на них агониста восстанавливается, чего не происходит при воздействии необратимых антагонистов.

Андроге́новый реце́птор, или андроге́нный рецептор, или NR3C4 — один из рецепторов стероидных гормонов, активируемый андрогенами — тестостероном или дигидротестостероном. Относится к подсемейству 3, группе С семейства ядерных рецепторов, способных непосредственно взаимодействовать с ядерной ДНК. Андрогеновый рецептор активируется при связывании с андрогенами в цитоплазме, а затем переносится в ядро.

Инсулиновый рецептор (IR) — трансмембранный рецептор, который активируется инсулином, IGF-I, IGF-II и принадлежит к большому классу тирозинкиназных рецепторов. Инсулиновый рецептор играет ключевую роль в регуляции гомеостаза глюкозы, функционального процесса, который при дегенеративных условиях может привести к ряду клинических проявлений, в том числе диабету и раку. Биохимически инсулиновый рецептор кодируется одним геном INSR, альтернативный сплайсинг которого в ходе транскрипции производит либо к IR-A-либо к IR-B-изоформам. Последующие посттрансляционные события каждой изоформы приводят к формированию протеолитически расщеплённых α и β-субъединиц, которые при объединении, в конечном счете способны к димеризации с получением дисульфидно связанного трансмембранного инсулинового рецептора массой ≈ 320 кДа.
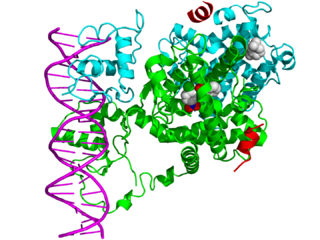
Ядерные рецепторы — это класс внутриклеточных белков, которые отвечают за восприятие стероидных и тиреоидных гормонов, а также некоторых других молекул. Уникальным свойством ядерных рецепторов, которое отличает их от других классов рецепторов, является их способность напрямую взаимодействовать с геномной ДНК и регулировать экспрессию соседних генов, тем самым контролируя развитие, гомеостаз и метаболизм организма. Следовательно, эти рецепторы классифицируются как факторы транскрипции. Регуляция экспрессии генов ядерными рецепторами обычно происходит только тогда, когда присутствует лиганд-молекула, которая влияет на поведение рецептора. Связывание лиганда с ядерным рецептором приводит к конформационному изменению рецептора, которое, в свою очередь, активирует рецептор, что приводит к усилению или понижению регуляции экспрессии гена.