Физиологические функции таламуса
Физиологические функции таламуса — это физиологические функции, выполняемые таламусом, отделом промежуточного мозга хордовых животных.
Таламус выполняет множество физиологических функций. Он может рассматриваться как своего рода «центральная релейно-трансформаторная станция» головного мозга, или как своеобразный хаб, куда стекается почти вся сенсорная и моторная информация (за исключением информации от органов обоняния) перед её передачей в кору больших полушарий головного мозга. Принято считать, что таламус действует как своеобразное реле или ретранслятор информации между различными подкорковыми областями и соответствующими областями коры больших полушарий головного мозга.[1] В частности, каждая из сенсорных систем, кроме обонятельной системы, включает в себя определённое таламическое ядро, которое принимает сенсорные сигналы от соответствующего первичного анализатора в стволе мозга, обрабатывает их и передаёт в соответствующую первичную область коры больших полушарий мозга.[2]
Ранее считалось, что таламус является всего лишь центральным «реле» или ретрансляторной станцией, которая просто передаёт различные сенсорные и моторные сигналы в кору больших полушарий головного мозга. Более поздние исследования показали, что функции таламуса гораздо сложнее, многообразнее и селективнее, и не сводятся лишь к простой ретрансляции информации от нижележащих подкорковых областей и структур головного мозга к коре больших полушарий, и что таламус также выполняет некую первичную обработку и фильтрацию поступающей в него сенсорной информации перед передачей её в кору больших полушарий.[3][2]
Сейчас полагают, что таламус не просто механически ретранслирует всю сенсорную и моторную информацию от органов чувств (вернее, от нижележащих подкорковых структур — первичных сенсорных анализаторов) в кору больших полушарий мозга, но и производит некую её первичную обработку и фильтрацию. Каждое из ядер таламуса, специализирующееся на первичной ретрансляции сенсорной информации того или иного типа в кору больших полушарий, получает сильные обратные связи от соответствующей зоны коры больших полушарий, модулирующие активность этого ядра и степень фильтрации им входящего потока информации.[2]
Роль таламуса в обработке сенсорной, моторной, висцеросенсорной и соматосенсорной информации
Различные ядра и области таламуса выполняют различные специфические функции. В частности, это относится ко многим сенсорным системам (системам органов чувств) таким как слуховая, зрительная, соматосенсорная (проприоцептивная), висцеросенсорная (интероцептивная) системы, система ощущения вкуса, за исключением обонятельной системы. Для каждой из этих систем существуют свои специализированные ядра таламуса, выполняющие функцию центральной релейной или ретрансляторной станции именно для этой системы. Изолированные локальные поражения этих ядер таламуса вызывают специфические нейросенсорные нарушения или дефициты в системе восприятия информации от соответствующих органов чувств.[2]
Так, например, для зрительной системы, вся входящая информация от сетчатки глаз передаётся в латеральное коленчатое тело, а уже оно, в свою очередь, направляет эту информацию, после её первичной обработки, в зрительную кору в затылочных долях коры больших полушарий мозга. Аналогично, медиальное коленчатое тело является центральным реле или ретрансляторной станцией для всей звуковой (слуховой) информации. Это ядро передаёт всю поступающую от нижнего холмика четверохолмия — структуры, расположенной в среднем мозге — слуховую и звуковую информацию, после её первичной обработки, в первичную слуховую кору. В свою очередь, вентральное заднее ядро таламуса является центральным реле для всей соматосенсорной, тактильной и проприоцептивной информации, поступающей от спинного мозга, и направляет её в первичную соматосенсорную кору.[2]
Роль таламуса в регуляции уровня сознания, цикла сон-бодрствование, концентрации внимания
Таламус также играет важную роль в регуляции уровня сознания, общего уровня возбуждения ЦНС, в регуляции концентрации внимания, смены состояний сна и бодрствования[4]. Ядра таламуса имеют множество сильных реципрокных (двусторонних) взаимных связей с корой больших полушарий головного мозга. Эти связи образуют кругообразно замкнутые таламо-кортико-таламические и кортико-таламо-кортикальные цепи, которые, как считается, связаны с регулированием уровня сознания, уровня возбуждения ЦНС, концентрации внимания, смены состояний сна и бодрствования. Повреждение таламуса может привести к летаргическому сну или перманентной (постоянной) коме или, наоборот, к упорной бессоннице[2].
По мнению известного британского нейрофизиолога А. Сета сознание зависит не от деятельности мозга в целом, а от коммуникации разных его частей между собой. В частности, от активности таламокортикальной системы, объединяющей кору головного мозга и таламус, соединенный с корой сложной сетью связей[5].
Роль таламуса в работе двигательной системы и систем языка и речи
Помимо передачи сенсорной, проприоцептивной, интероцептивной и моторной информации в кору больших полушарий, таламус также играет важную роль в интеграции работы и поддержании функционирования двигательной системы и системы языка и речи. Большая часть нейросхем таламуса, задействованных в регуляции работы этих сложных систем, вовлекает не одно, а несколько таламических ядер или групп ядер.[2]
Таламус также участвует во множестве нейронных информационных цепей, необходимых для управления двигательной подсистемой, и выполняет роль ключевого подкоркового центра регуляции движений, являясь подкорковым моторным центром «высшего порядка» по отношению к мозжечку и базальным ганглиям.[6] Благодаря исследованиям анатомии мозга приматов,[7] удалось выяснить характер многочисленных взаимосвязей ядер таламуса с мозжечком, базальными ганглиями и моторной (двигательной) корой. Это позволило выдвинуть предположение, что таламус выполняет функцию ключевого центра связи и передачи моторной информации по специализированным каналам от базальных ганглиев и мозжечка к моторной коре.[8][9] При исследовании антисаккадных движений глаз[10][11] у обезьян было обнаружено, что ядра таламуса участвуют в генерации антисаккадных движений глаз.[12]
Однако роль таламуса в регуляции функций нижележащих и более передне расположенных структур системы базальных ядер, в частности, нигростриарной и стрио-паллидарной систем, участвующих в двигательных актах, хотя и общепризнана, но всё ещё относительно плохо изучена. Роль таламуса в регуляции вестибулярной функции (функции равновесия) и функций ориентировки четверохолмия часто недооценивается или игнорируется, и также пока ещё плохо изучена.[2]
Роль таламуса в работе систем памяти и эмоций (лимбики и гиппокампа)
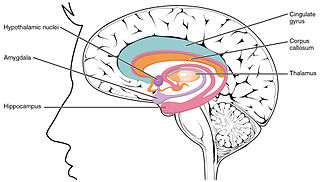
Передние ядра таламуса тесно функционально связаны с гиппокампом и структурами лимбической системы,[13] и часто рассматриваются как составная часть расширенной системы гиппокампа и лимбики.[14] Предоставляемая такой функциональной связью передних ядер таламуса с гиппокампом и лимбической системой возможность тесной интеграции предоставляемой гиппокампом пространственной и временной памяти, предоставляемой таламусом пространственно-временной сенсорной информации (например, информации о звуках, зрительных образах и других сенсорных ощущениях, сопровождавших то или иное событие, в привязке ко времени события и к конкретному месту в пространстве, где это событие происходило), и эмоциональной оценки события, предоставляемой лимбической системой — играет решающую роль в формировании интегрального воспоминания о картине того или иного эпизода во всей его целостности, вместе со всеми сопутствующими ему пространственно-временными метками, звуками, образами и другими сенсорными ощущениями, и его эмоциональной оценкой как положительного или отрицательного. Таким образом, тесные функциональные связи передних ядер таламуса с гиппокампом и структурами лимбической системы играют ключевую роль в формировании как человеческой эпизодической памяти, так и памяти событий у грызунов и других млекопитающих.[15][16]
Существует гипотеза о том, что связи определённых областей таламуса с определёнными областями мезио-темпоральной (средней височной) доли коры больших полушарий играют важную роль в дифференциации функционирования памяти пассивных воспоминаний и памяти узнавания знакомых мест, предметов и др., как у человека, так и у других млекопитающих.[17]
Примечания
- ↑ Michael S. Gazzaniga, Richard B. Ivry, George R. Mangun. Cognitive Neuroscience: The Biology of the Mind : [англ.]. — 4-е изд. — Нью-Йорк : W. W. Norton & Company, 2013. — 752 с. — ISBN 978-0393913484. — . — OCLC 908087478.
- ↑ 1 2 3 4 5 6 7 8 Darlene Susan Melchitzky, David A. Lewis. 1.2 Functional Neuroanatomy // Kaplan and Sadock's Comprehensive Textbook of Psychiatry : [англ.] : [арх. 29 сентября 2017] : in 2 vol. / под ред. Benjamin J. Sadock, Virginia A. Sadock, Pedro Ruiz. — 10-е изд. — Lippincott Williams & Wilkins, 2017. — Thalamus. — С. 158-170. — 12754 (эл.), 4997 (бум.) с. — ISBN 978-1451100471. — . — OCLC 949866139.
- ↑ Abigail W. Leonard. Your Brain Boots Up Like a Computer (англ.). livescience.com (17 августа 2006). Дата обращения: 27 сентября 2017. Архивировано 12 июля 2017 года.
- ↑ Mircea Steriade, Rodolfo R. Llinas. The functional states of the thalamus and the associated neuronal interplay : [англ.] : [арх. 16 сентября 2017] // Physiological Reviews. — 1988. — Т. 68, № 3 (July). — С. 649-742. — ISSN 1522-1210. — OCLC 114111014. — PMID 2839857.
- ↑ Сет, 2023, с. 51.
- ↑ E. V. Evarts, W. T. Thach. Motor Mechanisms of the CNS: Cerebrocerebellar Interrelations : [англ.] : [арх. 8 марта 2021] // Annual Review of Physiology. — 1969. — Т. 31 (March). — С. 451-498. — doi:10.1146/annurev.ph.31.030169.002315. — OCLC 4761161312. — PMID 4885774.
- ↑ Paul J. Orioli, Peter L. Strick. Cerebellar connections with the motor cortex and the arcuate premotor area: An analysis employing retrograde transneuronal transport of WGA-HRP : [англ.] : [арх. 28 сентября 2017] // The Journal of Comparative Neurology. — 1989. — Т. 288, № 4 (22 October). — С. 612-626. — ISSN 1096-9861. — doi:10.1002/cne.902880408. — OCLC 118356328. — PMID 2478593.
- ↑ C. Asanuma, W. T. Thach, E. G. Jones. Cytoarchitectonic delineation of the ventral lateral thalamic region in the monkey : [англ.] // Brain Research Reviews. — 1983. — Т. 5(286), № 3 (May). — С. 219-235. — ISSN 0006-8993. — doi:10.1016/0165-0173(83)90014-0. — OCLC 4929455787. — PMID 6850357.
- ↑ Kiyoshi Kurata. Activity Properties and Location of Neurons in the Motor Thalamus That Project to the Cortical Motor Areas in Monkeys : [англ.] : [арх. 28 сентября 2017] // Journal of Neurophysiology. — 2005. — Т. 94, № 1 (1 July). — С. 550-566. — ISSN 1522-1598. — doi:10.1152/jn.01034.2004. — OCLC 110603384. — PMID 15703228.
- ↑ Burkhart Fischer, Stefan Everling. The Antisaccade: A Review of Basic Research and Clinical Studies (англ.). Optomotorik.de. Фрайбург: Университет Фрайбурга. Дата обращения: 28 сентября 2017. Архивировано 16 сентября 2017 года.
- ↑ Stefan Everling, Burkhart Fischer. The antisaccade: a review of basic research and clinical studies : [англ.] // Neuropsychologia. — 1998. — Т. 36, № 9 (1 September). — ISSN 0028-3932. — doi:10.1016/S0028-3932(98)00020-7. — OCLC 4924629675. — PMID 9740362.
- ↑ Jun Kunimatsu, Masaki Tanaka. Roles of the Primate Motor Thalamus in the Generation of Antisaccades : [англ.] : [арх. 1 сентября 2017] // Journal of Neuroscience. — 2010. — Т. 30, № 14 (1 April). — С. 5108-5117. — ISSN 0270-6474. — doi:10.1523/JNEUROSCI.0406-10.2010. — OCLC 605093762. — PMID 20371831.
- ↑ Thor Stein, Chad Moritz, Michelle Quigley, Dietmar Cordes, Victor Haughton, Elizabeth Meyerand. Functional Connectivity in the Thalamus and Hippocampus Studied with Functional MR Imaging : [англ.] : [арх. 18 августа 2017] // American Journal of Neuroradiology. — 2000. — Т. 21, № 8 (September). — С. 1397-1401. — ISSN 0195-6108. — OCLC 199701670. — PMID 11003270.
- ↑ John P. Aggleton, Malcolm W. Brown. Episodic memory, amnesia, and the hippocampal–anterior thalamic axis : [англ.] : [арх. 28 сентября 2017] // Behavioral and Brain Sciences. — 1999. — Т. 22, № 3 (June). — С. 425-444; дискуссия на стр. 444-489. — ISSN 1469-1825. — doi:10.1017/S0140525X99002034. — OCLC 4669514763. — PMID 11301518.
- ↑ John P. Aggleton, Shane M. O’Mara, Seralynne D. Vann, Nick F. Wright, Marian Tsanov, Jonathan T. Erichsen. Hippocampal–anterior thalamic pathways for memory: uncovering a network of direct and indirect actions : [англ.] : [арх. 28 сентября 2017] // European Journal of Neuroscience. — 2010. — Т. 31, № 12 (14 June). — С. 2292–2307. — ISSN 1460-9568. — doi:10.1111/j.1460-9568.2010.07251.x. — OCLC 5151632719. — PMID 20550571. — PMC 2936113.
- ↑ Neil Burgess, Eleanor A Maguire, John O'Keefe. The Human Hippocampus and Spatial and Episodic Memory : [англ.] : [арх. 24 июля 2022] // Neuron. — 2002. — Т. 35, № 4 (15 August). — С. 625-641. — ISSN 0896-6273. — doi:10.1016/S0896-6273(02)00830-9. — OCLC 198675262. — PMID 12194864.
- ↑ Giovanni Augusto Carlesimo, Maria Giovanna Lombardi, Carlo Caltagirone. Vascular thalamic amnesia: A reappraisal : [англ.] // Neuropsychologia. — 2011. — Т. 49, № 5 (April). — С. 777-789. — ISSN 0028-3932. — doi:10.1016/j.neuropsychologia.2011.01.026. — OCLC 4803930095. — PMID 21255590.
Литература
- Сет, Анил. Быть собой. Новая теория сознания = Anil Seth. Being you: A New Science of Consciousness. — М.: Альпина нон-фикшн, 2023. — 400 с. — ISBN 978-5-00139-946-9.