Апопто́з — регулируемый процесс программируемой клеточной гибели, в результате которого клетка распадается на отдельные апоптотические тельца, ограниченные плазматической мембраной. Фрагменты погибшей клетки обычно очень быстро фагоцитируются макрофагами либо соседними клетками, минуя развитие воспалительной реакции. Морфологически регистрируемый процесс апоптоза продолжается 1—3 часа. Одной из основных функций апоптоза является уничтожение дефектных клеток. В многоклеточных организмах апоптоз к тому же задействован в процессах дифференциации и морфогенеза, в поддержании клеточного гомеостаза, в обеспечении важных аспектов развития и функционирования иммунной системы. Апоптоз наблюдается у всех эукариотов, начиная от одноклеточных простейших и вплоть до высших организмов. В программируемой смерти прокариотов участвуют функциональные аналоги эукариотических белков апоптоза.

Систе́ма комплеме́нта — комплекс защитных белков, постоянно присутствующих в крови. Это каскадная система протеолитических ферментов, предназначенная для гуморальной защиты организма от действия чужеродных агентов, она участвует в реализации иммунного ответа организма. Является важным компонентом как врождённого, так и приобретённого иммунитета. Выделяют три основных пути активации системы комплемента: классический, альтернативный и лектиновый. Для запуска классического пути комплемента необходима опсонизация чужеродной клетки антителами, а альтернативный и лектиновый пути могут активироваться в отсутствие антител. Поздние стадии у всех трёх путей активации системы комплемента одинаковы и включают образование мембраноатакующего комплекса, который нарушает целостность мембраны клетки-патогена и приводит к её гибели.
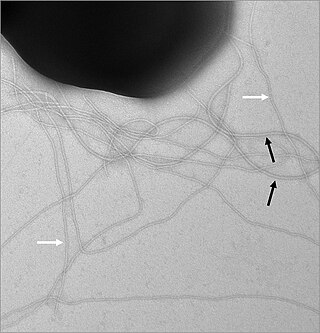
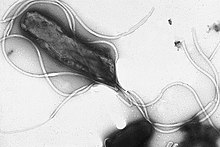
Жгу́тик — поверхностная структура, присутствующая у многих прокариотических и эукариотических клеток и служащая для их движения в жидкой среде или по поверхности твёрдых сред.
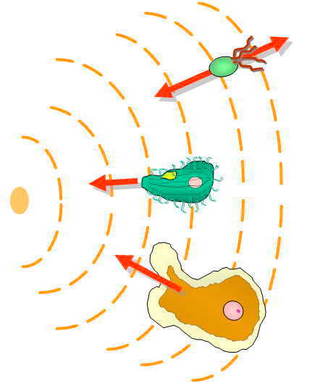
Хемота́ксис — двигательная реакция микроорганизмов на химический раздражитель.

Толл-подобные рецепторы — класс клеточных рецепторов с одним трансмембранным фрагментом, которые распознают консервативные структуры микроорганизмов и активируют клеточный иммунный ответ. Играют ключевую роль во врождённом иммунитете. Например, толл-подобный рецептор 4 узнаёт и связывается с консервативной структурой клеточной стенки грамотрицательных бактерий — липополисахаридом. Название получили благодаря сходству с белком, который кодируется открытым в 1985 году геном Toll у дрозофилы.
IRAK2 — цитозольная киназа, одна из четырёх сигнальных молекул, относящихся к группе IRAK. Участвует в передаче сигнала от рецептора интерлейкина-1 и некоторых толл-подобных рецепторов. Открыта в 1997 году по гомологии с IRAK1.

IRAK4 — белок, одна из четырёх цитозольных киназ группы сигнальных киназ IRAK, является важнейшей функциональной киназой своей группы. Участвует в передаче сигнала от рецептора интерлейкина-1 и некоторых толл-подобных рецепторов. Отсутствие или её недостаточность приводит к нарушению иммунного ответа на бактериальные инфекции. Открыта в 2002 году.
Протеинкина́зы — подкласс ферментов киназ (фосфотрансфераз). Протеинкиназы модифицируют другие белки путём фосфорилирования остатков аминокислот, имеющих гидроксильные группы или гетероциклической аминогруппы гистидина.
Иммунология насекомых — отрасль иммунологии, изучающая иммунитет насекомых.
TLR5 — мембранный белок, входящий в группу толл-подобных рецепторов, обеспечивающих функционирование врождённого иммунитета. TLR5, как и другие рецепторы этой группы, распознаёт патоген-связанные молекулярные структуры. Белок был обнаружен в 1998 году. Лигандом для TLR5 является флагеллин, бактериальный белок, главный компонент жгутиков, сходный у многих видов бактерий.
Рецепторы опознавания паттерна, или образ-распознающие рецепторы, — это белки, присутствующие на поверхности клеток иммунной системы и способные узнавать стандартные молекулярные структуры (паттерны), специфичные для больших групп патогенов. Их также называют рецепторами, опознающими патоген. По сравнению с системой адаптивного иммунитета, такие рецепторы и связанные с ними механизмы иммунной защиты являются эволюционно более древними.

DDR2 — мембранный белок, фермент из надсемейства рецепторных тирозинкиназ, продукт гена DDR2.
Трансформирующий фактор роста бета — белок, который контролирует пролиферацию, клеточную дифференцировку и другие функции в большинстве клеток. Участвует в иммунном ответе, раке, сердечно-сосудистых заболеваниях, сахарном диабете, синдроме Марфана, синдроме Лойеса-Дитса, болезни Паркинсона и синдроме приобретённого иммунодефицита (СПИД).
Сигнальные пути MAPK — группа мультифункциональных внутриклеточных сигнальных путей, содержащих одну из митоген-активируемых протеинкиназ и контролирующих транскрипцию генов, метаболизм, пролиферацию и подвижность клеток, апоптоз и другие процессы.

TrkB, тропомиозиновый тирозинкиназный рецептор — белок, кодируемый у человека геном NTRK2. В основном выступает в роли рецептора нейротропного фактора мозга (BDNF)
Отношения «ген против гена» были обнаружены Гарольдом Генри Флором, который работал с ржавчинными грибами, паразитирующими на льне. Флор показал, что наследование устойчивости у растения хозяина и наследование способности паразита вызывать заболевание (вирулентность) контролируется парами соответствующих друг другу генов. Гены растения, обуславливающие его устойчивость к патогену обозначаются — R. Комплементарные гены патогена изначально назывались генами авирулентности (Avr), поскольку они считались родственными генам устойчивости. Растения, образующие специфический продукт гена R, устойчивы к возбудителю, который производит соответствующий продукт гена Avr. Отношения «ген против гена» широко распространённый и являются очень важной особенностью устойчивости растений к патогенам. Например, отношения ген против гена хорошо изучены между латуком.
Бактериа́льные систе́мы секре́ции — белковые комплексы, расположенные в клеточной мембране бактерий и служащие для секреции различных белков. В частности, их используют патогенные бактерии для выделения факторов вирулентности. На основании состава, структуры и действия системы секреции делят на несколько типов. Наиболее фундаментальные различия наблюдаются между системами секреции грамположительных и грамотрицательных бактерий. Существует по меньшей мере шесть типов систем секреции, специфичных для грамотрицательных бактерий, четыре типа систем секреции уникальны для грамположительных бактерий, а два типа систем секреции имеются у обеих групп бактерий.

Интегральный мембранный белок NKG2-D II типа (NKG2-D type II integral membrane protein; CD314) — трансмембранный белок семейства CD94/NKG2 рецепторов лектина типа С, продукт гена человека KLRK1, локализованного NK-генном комплексе на 6-й хромосоме у мыши и 12-й хромосоме у человека. У человека белок экспрессирован на естественных киллерах, γδ-T-лимфоцитах и CD8+ αβ T-клетках и активированных макрофагах. NKG2D является рецептором к самоиндуцированным белкам-антигенам семейств MIC и RAET1/ULBP, которые появляются на поверхности клеток, находящихся в стрессовом состоянии, злокачественных клеток и инфицированных клеток.

Рецепторные тирозинкиназы, рецепторы с тирозинкиназной активностью — метаботропные клеточные рецепторы, представляют собой трансмембранные белки, которые состоят из каталитического внутриклеточного домена, участвующего в фосфорилировании субстратов (тирозинкиназа), трансмембранного и внешнего домена (связывающего) с высоким сродством к лигандам: гормонам (инсулин), цитокинам и полипептидным факторам роста. Из 90 уникальных генов тирозинкиназы, идентифицированных в геноме человека, 58 кодируют рецепторные тирозинкиназы.

MST1R — мембранный белок, рецепторная тирозинкиназа, продукт гена человека MST1R.