Фолликулярные дендритные клетки
Фолликулярные дендритные клетки (FDC)-это клетки иммунной системы, которые находятся в первичных и вторичных лимфатических фолликулах (лимфатических узлах) областей B-клеток лимфоидной ткани. В отличие от дендритных клеток (DC), FDC не происходят от костного мозга гемопоэтических стволовых клеток, а имеют Мезенхима происхождение. Возможные функции FDC включают: организацию клеток и микроархитектуры Лимфатическая система, захват антигена для поддержки B-клеток, содействие удалению мусора из герминальных центров и защиту от автоиммунных заболеваний. Заболевания, в которых могут участвовать FDC, включают первичные опухоли FDC, Воспаление, развитие Вирус иммунодефицита человека инфекции и невроинвазивный Почесуха овец.
Локализация и молекулярные маркеры
ФДК — немигрирующая популяция клеток, населяющая первичные и вторичные фолликулы. В клеточных зон лимфатических узлов, селезёнки и лимфоидной ткани, ассоциированной со слизистыми оболочками.
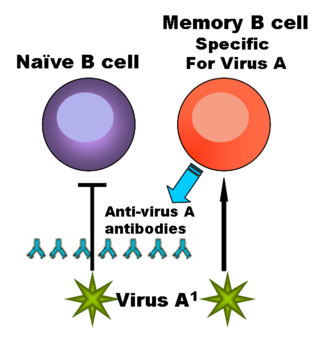
Благодаря тесному взаимодействию дендритных отростков, ФДК формируют стабильную сеть, которая составляет основу первичных и вторичных лимфоидных фолликулов[1][2]. Сеть ФДК не проникает в интерфолликулярную область и не затрагивает Т-клеточных зон лимфоидной ткани. Предположительно, смысл такой изоляции — в длительном сохранении антигенов, опсонизированных и представленных на поверхности ФДК, для поддержания популяций В-лимфоцитов памяти.
ФДК в большом количестве экспрессируют рецепторы комплемента CR1 и CR2 (CD35 и CD21), а также Fc-рецептор FcγRIIb (CD32). Молекулярными маркерами ФДК так же являются: FDC-M1, FDC-M2 и C4. ФДК не экспрессируют на своей поверхности молекулы второго класса главного комплекса гистосовместимости (MHC). На них также практически отсутствуют рецепторы опознавания паттерна (PRR), поэтому ФДК не способны самостоятельно распознавать и презентовать антиген клеткам иммунной системы, до тех пор, пока он не опсонизирован антителами или молекулами комплемента.
Развитие
Фолликулярные дендритные клетки развиваются из мигрирующих мезенхимальных клеток-предшественников[3]. На мышах с острым общим иммунодефицитом (англ. severe combined immunodeficiency) была продемонстрирована возможность переноса мезенхимальных клеточных предшественников ФДК реципиенту костного мозга. ФДК у такого реципиента развиваются как из собственных клеток-предшественников, так и из клеток-предшественников донора[4]. Воздействие лимфоцитов на предшественники ФДК посредством синтеза фактора некроза опухоли альфа (ФНО-α; TNF-α) и лимфотоксина бета (LT-β) играет решающую роль в развитии и поддержании популяции ФДК[]. Для ФНО-α на поверхности прекурсоров ФДК экспессируется рецептор TNF-RI, лимфотоксин же взаимодействует с рецептором LT-β[]. У мышей, с отсутствием B-лимфоцитов и нарушенным синтезом ФНО-α и лимфотоксина, фолликулярные дендритические клетки не развиваются и не образуют сети в фолликулах[].
Функции
Организация лимфатической микроструктуры
В нормальной лимфатической ткани неактивные B-лимфоциты мигрируют сквозь сети фолликулярных дендритных клеток, тогда как движение активированных антигеном B-клеток в ФДК сети замедляется, и они вступают в стадию клональной экспансии, формируя центры размножения, или герминативные центры (ГЦ). Фолликулярные дендритные клетки в большом количестве синтезируют CXCL13 — хемоаттрактант, который привлекает В лимфоциты и организует лимфоидную ткань[5].
Презентация опсонизированных антигенов и поддержание пула B-клеток памяти
Фолликулярные дендритные клетки удерживают опсонизированные комплементом или антителами антигены с помощью рецепторов CR1, CR2 и FcγRIIb. Только те B-клетки, которые способны связаться с антигеном, представленным на поверхности ФДК, выживают и сохраняют способность производить антитела, становясь B-лимфоцитами памяти. Все прочие активированные B-лимфоциты получают сигнал к инициации апоптоза[2].
Участие в уничтожении апоптотических телец
Поскольку ФДК способствуют запуску апоптоза в активированных В-лимфоцитах, не связавшихся с антигеном, на поверхности ФДК, а также в B-клетках, проявляющих признаки аутоагрессии, количество апоптотических телец в ФДК сети герминативного центра высоко. Фолликулярные дендритические клетки, секретируют фактор Mfge8, который связывает апоптотические клетки с макрофагами, ускоряя поглощение апоптотических телец и делая более эффективным устранение молекулярного «мусора» из герминативного центра[6][7].
Предотвращение аутоиммунных заболеваний
Фактор Mfge8, продуцируемый в лимфатической ткани преимущественно ФДК ускоряет поглощение апоптотических телец макрофагами. Отсутствие этого фактора у мышей приводит к состоянию, напоминающему системную красную волчанку. Кроме того, мыши, у которых нарушена продукция лимфотоксина, необходимого для стимуляции ФДК, или отсутствует рецептор к нему, демонстрируют тенденцию к образованию массивных лимфоцитарных инфильтратов, которые, предположительно, могут запускать аутоиммунный процесс. Таким образом, вероятно, что ФДК, участвуют в подавлении аутоиммунных реакций[6].
Взаимодействия с B-клетками
Неконгенированные (не антигенспецифичные) В-клетки играют важную роль в транспорте антигенов к FDC. Они захватывают иммунные комплексы CR1/2-зависимым способом либо непосредственно из лимфы, либо из макрофагов и перемещаются в лимфоидную ткань, где переносят опсонизированный комплементом антиген в FDCS.
FDC, в свою очередь, привлекают В-клетки хемоаттрактантом CXCL13. В-клетки, лишенные CXCR5, рецептора для CXCL13, все еще проникают в белую пульпу, но неправильно локализованы и дезорганизованы. Для формирования фолликулярных структур FDCS необходимо стимулировать лимфотоксином (LT), медиатором, вырабатываемым В-клетками. Стимуляция CXCR5 В В-клетках усиливает выработку LT, что приводит к активации FDCs и стимулирует дальнейшую секрецию CXCL13, создавая таким образом положительный цикл обратной связи. Это приводит к образованию зародышевых центров (GCS), где активированные антигеном В-клетки попадают в ловушку для прохождения соматической мутации, положительного и отрицательного отбора, переключения изотипа и дифференцировки в высокоаффинные плазматические клетки и В-клетки памяти. Адгезия между FDCS и В-клетками опосредуется молекулами ICAM–1 (CD54)-LFA–1 (CD11a) и VCAM-VLA-4. Активированные В-клетки с низким сродством к антигену, захваченные на поверхности FDCS, а также аутореактивные В-клетки подвергаются апоптозу,тогда как В-клетки, связанные с FDCS через антигенный комплекс, выживают благодаря блокировке апоптоза, вызванной взаимодействием с FDCs.
Болезни
Описаны редкие первичные FDC-опухоли. Эти саркомы часто поражают лимфоидные ткани, но в ряде случаев опухоль была обнаружена в печени, желчных протоках, поджелудочной железе, щитовидной железе, носоглотке, небе, подслизистой оболочке желудка или двенадцатиперстной кишки. При ряде хронических воспалительных состояний клетки, продуцирующие хемокин CXCL13 и несущие такие маркеры FDCs, как VCAM-1 и CD21, наблюдались в совершенно неожиданных местах, включая синовиальную ткань пациентов с ревматоидным артритом (РА), слюнные железы пациентов с синдромом Шегрена и кожу пациентов с псевдобелкоклеточной лимфомой. Фолликулярные дендритные клетки участвуют в развитии инфекции ВИЧ-1, как обеспечивая убежище для ВИЧ-1, так и стимулируя репликацию ВИЧ-1 в соседних инфицированных моноцитарных клетках с помощью юкстакринового сигнального механизма.Также есть некоторые доказательства того, что FDCS могут способствовать репликации прионов и нейроинвазии при нейроинвазивной скрепи.
Примечания
- ↑ Male D., Brostoff J., Roth D., Roitt I. (неопр.). — 2007. — ISBN 9780323033992.
- ↑ 1 2 Banchereau J., Steinman R. M. Dendritic cells and the control of immunity (англ.) // Nature. — 1998. — Vol. 392. — P. 245—252. — doi:10.1038/32588. — PMID 9521319.
- ↑ van Nierop K., de Groot C. Human follicular dendritic cells: function, origin and development (англ.) // Semin Immunol : journal. — 2002. — Vol. 14, no. 4. — P. 251—257. — doi:10.1016/S1044-5323(02)00057-X. — PMID 12163300.
- ↑ Kapasi Z. F., Qin D., Kerr W. G., Kosco-Vilbois M. H., Shultz L. D., Tew J. G., Szakal A. K. Follicular dendritic cell (FDC) precursors in primary lymphoid tissues (англ.) // The Journal of Immunology : journal. — 1998. — Vol. 160, no. 3. — P. 1078—1084. — PMID 9570519.
- ↑ Cyster J. G. B cell follicles and antigen encounters of the third kind (англ.) // Nat Immunol. : journal. — 2010. — Vol. 11, no. 11. — P. 989—996. — doi:10.1038/ni.1946. — PMID 20959804.
- ↑ 1 2 Aguzzi A., Krautler NJ. Characterizing follicular dendritic cells: A progress report (англ.) // Eur J Immunol. : journal. — 2010. — Vol. 40, no. 8. — P. 2134—2138. — doi:10.1002/eji.201040765. — PMID 20853499.
- ↑ Kranich J., Krautler N. J., Heinen E., Polymenidou M., Bridel C., Schildknecht A., Huber C., Kosco-Vilbois M. H., Zinkernagel R., Miele G., Aguzzi A. Follicular dendritic cells control engulfment of apoptotic bodies by secreting Mfge8 (англ.) // Journal of Experimental Medicine[англ.] : journal. — Rockefeller University Press[англ.], 2008. — Vol. 205, no. 6. — doi:10.1084/jem.20071019. — PMID 18490487.