
Фотоси́нтез — сложный химический процесс преобразования энергии видимого света в энергию химических связей органических веществ при участии фотосинтетических пигментов.

Тилакоиды — ограниченные мембраной компартменты внутри хлоропластов и цианобактерий. В тилакоидах происходят светозависимые реакции фотосинтеза. Слово «тилакоид» происходит от греческого слова θύλακος, означающего «мешочек». Тилакоиды состоят из мембраны, окружающей просвет тилакоида. Тилакоиды хлоропластов часто имеют структуру, напоминающую стопку дисков. Эти стопки называют гранами. Граны соединены межграновыми или строматическими тилакоидами (ламеллами) в единое функциональное пространство.

Хлоросо́мы — обогащённые липидами везикулы зелёных серныx бактерий и нитчатых аноксигенных фототрофных бактерий, локализованные в цитоплазме и связанные с клеточной мембраной кристаллической базальной пластинкой. Внутри хлоросомы находятся пучки палочковидных структур, содержащих молекулы бактериохлорофиллов c, d или e. Таким образом, в хлоросомах собраны светособирающие системы.

Зелёные серобактерии (лат. Chlorobiaceae) — семейство облигатно анаэробных (более строгих, чем пурпурные бактерии, в присутствии O2 не растут) фотолитоавтотрофных грамотрицательных бактерий, использующих сероводород (H2S), водород (H2) и элементарную серу (S0) в качестве доноров электронов. По происхождению они принадлежат к надтипу Bacteroidetes-Chlorobi, однако неоднородны и потому их классифицируют как отдельный тип.

Феофитин — химическое соединение, служащее одним из первых акцепторов электрона в цепи переноса электронов в реакционном центре фотосистемы II (ФСII) у растений, и в реакционном центре (РЦ) пурпурных бактерий. Как в реакционном центре ФСII, так и бактерий (P870), возбуждённые электроны передаются от воды на феофитин, который затем передаёт их хинону (QA). В целом механизмы, функции и задачи молекулы феофитина аналогичны друг другу в обеих транспортных цепях.
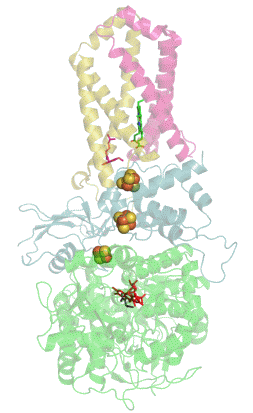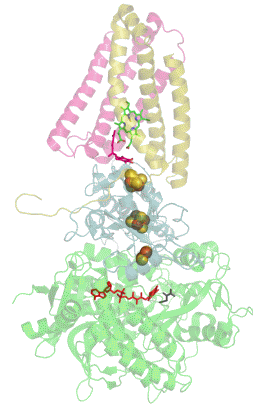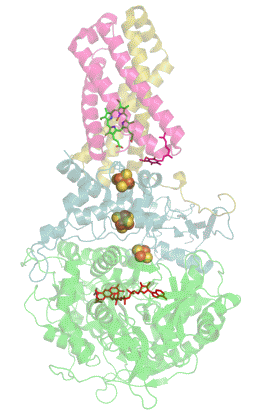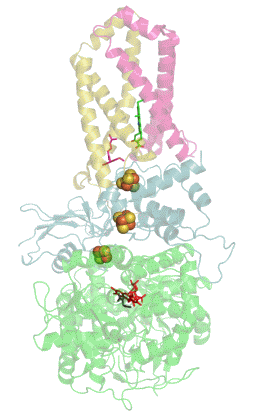
Сукцинатдегидрогеназа или сукцинат-убихинон-оксидорекдуктаза, также известная как комплекс II — белковый комплекс, расположенный во внутренней мембране митохондрий и мембранах многих прокариотических организмов. Одновременно участвует в цикле трикарбоновых кислот и дыхательной цепи переноса электронов.
Электрон-транспортная цепь фотосинтеза — последовательность переносчиков электронов, расположенных на белках фотосинтетических мембран и осуществляющих фотоиндуцированный транспорт электронов, сопряжённый с трансмембранным переносом протонов против электрохимического градиента.
Ферредокси́ны — группа небольших растворимых белков, содержащих железосерные кластеры и являющихся подвижными переносчиками электронов в ряде метаболических процессов. Обычно они переносят один или два электрона за счёт изменения окисленности атомов железа.

Светособирающие комплексы — пигмент-белковые комплексы фотосинтезирующих организмов, локализованные в фотосинтетических мембранах и выполняющие функцию первичного поглощения квантов света с последующей миграцией энергии возбуждения к реакционным центрам фотосистем. Также они обеспечивают тонкую настройку фотосинтетического аппарата и участвуют в его защите от фотоповреждений.

Фотосисте́ма I, или пластоциани́н-ферредокси́н-оксидоредукта́за — второй функциональный комплекс электрон-транспортной цепи (ЭТЦ) хлоропластов. Он принимает электрон от пластоцианина и, поглощая световую энергию, формирует сильный восстановитель П700, способный через цепь переносчиков электронов осуществить восстановление НАДФ+. Таким образом, при участии ФСI синтезируется источник электронов (НАДФН) для последующих реакций восстановления углерода в хлоропластах в цикле Кальвина. Кроме того, ФСI может осуществлять циклический транспорт электронов, сопряжённый с синтезом АТФ, обеспечивая дополнительный синтез АТФ в хлоропластах.

Реакционный центр — комплекс белков, пигментов и других кофакторов, взаимодействие которых обеспечивает реакцию превращения энергии света в химическую при фотосинтезе. Реакционный центр получает энергию или через непосредственное возбуждение одной из своих молекул или через перенос энергии от светособирающих комплексов, что даёт начало цепочке химических реакций, происходящей на связанных белками кофакторах. Эти кофакторы — светопоглощающие молекулы такие как хлорофилл, феофитин и хиноны. Энергия фотона используется для поднятия электрона на более высокий энергетический уровень. Запасённая таким образом свободная энергия идёт на восстановление цепочки акцепторов электрона с более высоким редокс-потенциалом.

Фотосисте́ма II (втора́я фотосисте́ма, фотосисте́ма два, ФСII), или H2O-пластохиноноксидоредуктаза — первый функциональный комплекс электрон-транспортной цепи (ЭТЦ) хлоропластов. Он расположен в мембранах тилакоидов всех растений, водорослей и цианобактерий. Поглощая энергию света в ходе первичных фотохимических реакций, он формирует сильный окислитель — димер хлорофилла a (П680+), который через цепь окислительно-восстановительных реакций способен вызвать окисление воды.

Цитохро́м-b6f-ко́мплекс, или пластохинолпластоцианинредукта́за — мультибелковый комплекс, который осуществляет окисление пластохинолов и восстановление белка пластоцианина, обеспечивая, таким образом, транспорт электронов между реакционными центрами фотосистемы I (ФСI) и фотосистемы II (ФСII). Он восстанавливает маленький водорастворимый белок пластоцианин, который переносит электрон к ФСII. Аналогичную реакцию катализирует цитохром-bc1-комплекс электрон-транспортной цепи митохондрий. Цитохром-b6f-комплекс присутствует в тилакоидной мембране хлоропластов растений, водорослей и цианобактерий. Он функционально объединяет две фотосистемы в единую цепь переноса электронов от воды к НАДФ+, то есть является участником нециклического потока электронов. Кроме того, цитохромный комплекс вовлечён в циклический транспорт электронов, осуществляемый фотосистемой I.
Нефотохимическое тушение — механизм защиты фотосинтетического аппарата от света высокой интенсивности, используемый растениями и водорослями. Суть процесса заключается в поглощении избыточной энергии (тушении) синглетного возбужденного хлорофилла молекулой-акцептором с последующим переходом этой молекулы в основное энергетическое состояние при помощи усиленной внутренней конверсии. Благодаря внутренней конверсии избыточная энергия возбуждения рассеивается в виде тепла, то есть расходуется на молекулярные колебания. Нефотохимическое тушение есть почти у всех фотосинтезирующих эукариот и цианобактерий. Оно помогает регулировать и защищать фотосинтетический аппарат в условиях, когда поглощается больше света, чем может быть непосредственно использовано в фотосинтезе.

Фотофосфорили́рование — процесс синтеза АТФ из АДФ за счёт энергии света. Как и в случае окислительного фосфорилирования, энергия света расходуется на создание протонного градиента на мембране тилакоидов или клеточной мембране бактерии, который затем используется АТФ-синтазой. Фотофосфорилирование — очень древняя форма фотосинтеза, которая есть у всех фототрофных эукариот, бактерий и архей. Различают два типа фосфорилирования — циклическое, сопряжённое с циклическим потоком электронов в электрон-транспортной цепи, и нециклическое, сопряжённое с прямым потоком электронов от H2O к НАДФ+ в случае эукариот или другого донора электрона в случае бактерий, например, H2S. Как разновидность нециклического типа выделяют псевдоциклическое фотофосфорилирование, при котором акцептором электронов служит кислород.

Фотоингибирование — индуцируемое светом снижение фотосинтетической активности растений, водорослей или цианобактерии. Фотосистема II (ФСII) более чувствительна к свету, чем другие фотосинтетические машины, поэтому большинство исследователей определяют термин как светоиндуцируемое повреждения ФСII. В живых организмах, поврежденные избыточным освещением фотосистемы II постоянно репарируются за счёт деградации и синтеза белка D1 фотосинтетического реакционного центра ФСII. Понятие фотоингибирование также используется в более широком смысле, в значении «динамическое фотоингибирование», чтобы описать все реакции, которые приводят к снижению эффективности фотосинтеза, когда растения подвергаются воздействию света.
П680 (P680, пигмент 680) или первичный донор фотосистемы II — димер из двух молекул хлорофилла а, П1 и П2, которые также называют специальной парой. Вместе две эти молекулы образуют экситонный димер, то есть они функционально представляют собой единую систему и при возбуждении ведут себя как одна молекула. Максимум поглощения энергии света такой специальной пары приходится на длину волны λ = 680 нм. Первичный донор возбуждается, поглощая фотоны с подходящей длиной волны или через перенос энергии возбуждения от других хлорофиллов фотосистемы II. П680 поглощает квант света и переходит в фотовозбуждённое состояние, в результате чего один из его электронов переходит на более высокий энергетический уровень — с основного подуровня S0 на первый синглетный подуровень S1. Этот электрон отрывается от специальной пары и захватывается первичным акцептором электрона, феофитином, который расположен внутри фотосистемы II рядом с П680. Процесс отщепления электрона от специальной пары и переходе его на феофитин с образованием радикальной пары носит название разделения зарядов. Окисленный П680+ восстанавливается, захватывая электрон от водоокисляющего комплекса фотосистемы II.
Цитохром b559 — гетеродимерный белок, состоящий из одной альфа (PsbE) и одной бета (PsbF) субъединицы, между которыми расположен гем; важный компонент фотосистемы II.
НАДН-дегидрогена́зный ко́мплекс хлоропла́стов — мультибелковый комплекс электронотранспортной цепи фотосинтеза, расположенный в тилакоидной мембране пластид высших растений и водорослей. Комплекс окисляет ферредоксин и восстанавливает молекулы пластохинона, которые высвобождаются в мембрану. При этом энергия окисленного восстановительного эквивалента расходуется на перенос протонов из стромы хлоропласта в люмен тилакоида с образованием протонного градиента. Показано большее сходство НАДН-дегидрогеназного комплекса хлоропластов с цианобактериальным НАДН дегидрогеназным комплексом (NDH-1), чем с митохондриальным комплексом I.
Ферредоксин-НАДФ+-редуктаза, сокращенно ФНР, фермент из класса оксидоредуктаз, катализирующий реакцию восстановления НАДФ+, используя в качестве донора электронов ферредоксин.
