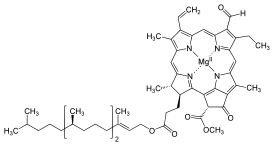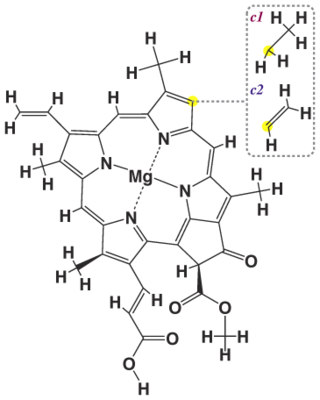
Хлорофи́лл — зелёный пигмент, окрашивающий хлоропласты растений в зелёный цвет. При его участии происходит фотосинтез. По химическому строению хлорофиллы — магниевые комплексы различных тетрапирролов. Хлорофиллы имеют порфириновое строение и близки гему. Хлорофилл зарегистрирован в качестве пищевой добавки Е140.

Фотоси́нтез — сложный химический процесс преобразования энергии видимого света в энергию химических связей органических веществ при участии фотосинтетических пигментов.

Тилакоиды — ограниченные мембраной компартменты внутри хлоропластов и цианобактерий. В тилакоидах происходят светозависимые реакции фотосинтеза. Слово «тилакоид» происходит от греческого слова θύλακος, означающего «мешочек». Тилакоиды состоят из мембраны, окружающей просвет тилакоида. Тилакоиды хлоропластов часто имеют структуру, напоминающую стопку дисков. Эти стопки называют гранами. Граны соединены межграновыми или строматическими тилакоидами (ламеллами) в единое функциональное пространство.

Фикобили́ны — тетрапиррольные пигменты (билины) красных водорослей, криптофит и цианобактерий. Фикобилины являются хромофорной группой фикобилипротеинов — кислых водорастворимых глобулярных хромопротеинов светособирающего комплекса водорослей. Отдельными молекулами фикобилины, как правило, не представлены, а образуют комплексы с белками — фикобилипротеиды (хромопротеиды).

Фикобилисо́мы — светособирающие органеллы для фотосистемы II у цианобактерий, красных водорослей и глаукофитов. Стандартные фикобилисомы отсутствуют у криптофитовых и тех представителей прохлорофитовых, у которых имеются фикобилипротеины. У криптофитовых фикобилипротеины находятся во внутритилакоидном пространстве.

Хлорофилл c1 — форма хлорофилла. Является вспомогательным пигментом, придаёт золотистый или коричневатый цвет. Помогает собирать энергию и передаёт её на светособирающую антенну в реакционный центр. Хлорофилл c необычен, поскольку у него нет изопреноидного хвоста, а пиррол D не окислен. Всё это нехарактерно для большинства хлорофиллов, обнаруженных в водорослях и растениях. Хлорофилл c был обнаружен в морских водорослях, таких как диатомовые водоросли. Максимумы поглощения: 444, 577, 626 нм и 447, 579, 629 нм в диэтиловом эфире и ацетоне соответственно.

Хлорофилл d — один из хлорофиллов, 3-дезвинил-3-формил-хлорофилл а. В растворах органических растворителей имеет красный максимум поглощения в области 690—697 нм, сдвинутый на 30 нм в длинноволновую сторону по сравнению с хлорофиллом a, а в живых организмах — 710 нм, то есть уже на границе инфракрасной области.

Хлорофи́лл a — особая форма хлорофилла, используемая для оксигенного фотосинтеза. Сильнее всего поглощает свет в фиолетово-голубой и оранжево-красной части спектра. Этот пигмент жизненно необходим для фотосинтеза в клетках эукариот, цианобактерий и прохлорофитов из-за своей способности отдавать возбуждённые электроны в электрон-транспортную цепь. Хлорофилл a также является частью антенного комплекса и передаёт резонансную энергию, которая затем поступает в реакционный центр, где расположены специальные хлорофиллы P680 и P700.

Протохлорофиллид, или моновинил протохлорофиллид, это непосредственный предшественник хлорофилла a с отсутствующим фитольным хвостом. В отличие от хлорофилла, протохлорофиллид имеет сильную флюоресценцию; мутанты, накапливающие его в своих тканях, светятся красным, если облучить их синим светом. У цветковых растений, реакция превращения протохлорофиллида в хлорофилл является светозависимой, и такие растения становятся белыми (хлорозными) если выращивать их в темноте. В отличие от них голосеменные, водоросли, и фотосинтезирующие бактерии используют другой, не зависящий от света фермент, и вырастают зелёными даже в темноте.

Светособирающие комплексы — пигмент-белковые комплексы фотосинтезирующих организмов, локализованные в фотосинтетических мембранах и выполняющие функцию первичного поглощения квантов света с последующей миграцией энергии возбуждения к реакционным центрам фотосистем. Также они обеспечивают тонкую настройку фотосинтетического аппарата и участвуют в его защите от фотоповреждений.

Фотосисте́ма I, или пластоциани́н-ферредокси́н-оксидоредукта́за — второй функциональный комплекс электрон-транспортной цепи (ЭТЦ) хлоропластов. Он принимает электрон от пластоцианина и, поглощая световую энергию, формирует сильный восстановитель П700, способный через цепь переносчиков электронов осуществить восстановление НАДФ+. Таким образом, при участии ФСI синтезируется источник электронов (НАДФН) для последующих реакций восстановления углерода в хлоропластах в цикле Кальвина. Кроме того, ФСI может осуществлять циклический транспорт электронов, сопряжённый с синтезом АТФ, обеспечивая дополнительный синтез АТФ в хлоропластах.

Реакционный центр — комплекс белков, пигментов и других кофакторов, взаимодействие которых обеспечивает реакцию превращения энергии света в химическую при фотосинтезе. Реакционный центр получает энергию или через непосредственное возбуждение одной из своих молекул или через перенос энергии от светособирающих комплексов, что даёт начало цепочке химических реакций, происходящей на связанных белками кофакторах. Эти кофакторы — светопоглощающие молекулы такие как хлорофилл, феофитин и хиноны. Энергия фотона используется для поднятия электрона на более высокий энергетический уровень. Запасённая таким образом свободная энергия идёт на восстановление цепочки акцепторов электрона с более высоким редокс-потенциалом.

Фотосисте́ма II (втора́я фотосисте́ма, фотосисте́ма два, ФСII), или H2O-пластохиноноксидоредуктаза — первый функциональный комплекс электрон-транспортной цепи (ЭТЦ) хлоропластов. Он расположен в мембранах тилакоидов всех растений, водорослей и цианобактерий. Поглощая энергию света в ходе первичных фотохимических реакций, он формирует сильный окислитель — димер хлорофилла a (П680+), который через цепь окислительно-восстановительных реакций способен вызвать окисление воды.
Вспомогательные пигменты — светопоглощающие соединения фотосинтезирующих организмов, которые работают в сочетании с хлорофиллом а, выполняя светособирающую или светозащитную функции. К ним относятся различные формы хлорофилла, например хлорофилл b в зеленых водорослях и светособирающих антеннах высших растений или хлорофиллы с или d у других водорослей. Кроме того есть много других вспомогательных пигментов таких как каротиноиды или фикобилипротеины, которые также поглощают свет и передают его энергию на главный пигмент фотосистемы. Некоторые из этих вспомогательных пигментов, в частности каротиноиды, служат для поглощения и рассеивания избыточной энергии света и являются антиоксидантами.
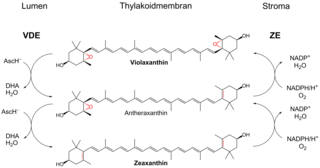
Ксантофилловый цикл, или виолаксантиновый цикл, в случае высших растений, выполняет функцию защиты фотосинтетического аппарата от избытка энергии при повышенной инсоляции. Он позволяет избежать фотоингибирования, значительно увеличив нефотохимическое тушение. Цикл включает в себя ферментативные взаимопревращения между виолаксантином и зеаксантином.
Нефотохимическое тушение — механизм защиты фотосинтетического аппарата от света высокой интенсивности, используемый растениями и водорослями. Суть процесса заключается в поглощении избыточной энергии (тушении) синглетного возбужденного хлорофилла молекулой-акцептором с последующим переходом этой молекулы в основное энергетическое состояние при помощи усиленной внутренней конверсии. Благодаря внутренней конверсии избыточная энергия возбуждения рассеивается в виде тепла, то есть расходуется на молекулярные колебания. Нефотохимическое тушение есть почти у всех фотосинтезирующих эукариот и цианобактерий. Оно помогает регулировать и защищать фотосинтетический аппарат в условиях, когда поглощается больше света, чем может быть непосредственно использовано в фотосинтезе.

Фотосистема представляет собой функциональную и структурную единицу белковых комплексов, которые осуществляют первичные фотохимические реакции фотосинтеза: поглощение света, преобразование энергии и перенос электронов. Фотосистемы находятся в тилакоидной мембраны растений, водорослей и цианобактерий, либо в цитоплазматической мембране фотосинтезирующих бактерий. В общем случае все фотосистемы подразделяют на два типа — подобные фотосистеме II и подобные фотосистеме I.

Фотоингибирование — индуцируемое светом снижение фотосинтетической активности растений, водорослей или цианобактерии. Фотосистема II (ФСII) более чувствительна к свету, чем другие фотосинтетические машины, поэтому большинство исследователей определяют термин как светоиндуцируемое повреждения ФСII. В живых организмах, поврежденные избыточным освещением фотосистемы II постоянно репарируются за счёт деградации и синтеза белка D1 фотосинтетического реакционного центра ФСII. Понятие фотоингибирование также используется в более широком смысле, в значении «динамическое фотоингибирование», чтобы описать все реакции, которые приводят к снижению эффективности фотосинтеза, когда растения подвергаются воздействию света.
Эффект Каутского — явление изменения свечения хлорофилла, которое возникает при освещении предварительно адаптированного к темноте листа растения. Этот феномен был открыт Х. Каутским и А. Хиршем в 1931 году.

Prochlorococcus marinus (лат.) — вид крошечных фотосинтезирующих цианобактерий из порядка Synechococcales, типовой и единственный в роде Prochlorococcus. Систематики не пришли к единому мнению, какому семейству отнести это род: к Synechococcaceae Komárek and Anagnostidis 1995 или к Prochloraceae R.A.Lewin.