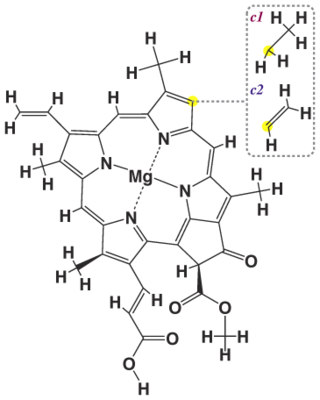
Хлорофи́лл — зелёный пигмент, окрашивающий хлоропласты растений в зелёный цвет. При его участии происходит фотосинтез. По химическому строению хлорофиллы — магниевые комплексы различных тетрапирролов. Хлорофиллы имеют порфириновое строение и близки гему. Хлорофилл зарегистрирован в качестве пищевой добавки Е140.

Тилакоиды — ограниченные мембраной компартменты внутри хлоропластов и цианобактерий. В тилакоидах происходят светозависимые реакции фотосинтеза. Слово «тилакоид» происходит от греческого слова θύλακος, означающего «мешочек». Тилакоиды состоят из мембраны, окружающей просвет тилакоида. Тилакоиды хлоропластов часто имеют структуру, напоминающую стопку дисков. Эти стопки называют гранами. Граны соединены межграновыми или строматическими тилакоидами (ламеллами) в единое функциональное пространство.

Страменопилы, страминопилы, или гетероконтные, разножгутиковые организмы (лат. Heterokonta), — большая группа одноклеточных, колониальных и многоклеточных эукариотических организмов. Для фотосинтезирующих представителей характерно наличие хлорофиллов a, c1, c2, c3, а также фукоксантина или вашериоксантина. Длинный жгутик в норме покрыт трёхчастными мастигонемами (ретронемами).

Хлорофилл c1 — форма хлорофилла. Является вспомогательным пигментом, придаёт золотистый или коричневатый цвет. Помогает собирать энергию и передаёт её на светособирающую антенну в реакционный центр. Хлорофилл c необычен, поскольку у него нет изопреноидного хвоста, а пиррол D не окислен. Всё это нехарактерно для большинства хлорофиллов, обнаруженных в водорослях и растениях. Хлорофилл c был обнаружен в морских водорослях, таких как диатомовые водоросли. Максимумы поглощения: 444, 577, 626 нм и 447, 579, 629 нм в диэтиловом эфире и ацетоне соответственно.

Хлорофилл c2 — наиболее распространенная форма хлорофиллов с. Присутствует в качестве дополнительного пигмента почти у всех фотосинтезирующих хромальвеолят ("хромист"), за исключением эустигматофитовых и синурофитовых.

Хлорофилл b — форма хлорофилла, один из вспомогательных пигментов фотосинтеза у высших растений, зеленых водорослей и эвгленовых, а также у цианобактерий группы прохлорофит.

Хлорофилл f — форма хлорофилла, обнаруженная в 2010 году в строматолитах западно-австралийского залива Шарк. От других форм хлорофилла он отличается тем, что его спектр поглощения сильно смещён в длинноволновую часть красной области. Открытие совершила группа учёных из Сиднейского университета под руководством доктора Мин Чена, и это первое обнаружение новой формы хлорофилла за последние 60 лет.

Хлорофилл d — один из хлорофиллов, 3-дезвинил-3-формил-хлорофилл а. В растворах органических растворителей имеет красный максимум поглощения в области 690—697 нм, сдвинутый на 30 нм в длинноволновую сторону по сравнению с хлорофиллом a, а в живых организмах — 710 нм, то есть уже на границе инфракрасной области.

Хлорофи́лл a — особая форма хлорофилла, используемая для оксигенного фотосинтеза. Сильнее всего поглощает свет в фиолетово-голубой и оранжево-красной части спектра. Этот пигмент жизненно необходим для фотосинтеза в клетках эукариот, цианобактерий и прохлорофитов из-за своей способности отдавать возбуждённые электроны в электрон-транспортную цепь. Хлорофилл a также является частью антенного комплекса и передаёт резонансную энергию, которая затем поступает в реакционный центр, где расположены специальные хлорофиллы P680 и P700.

Протохлорофиллид, или моновинил протохлорофиллид, это непосредственный предшественник хлорофилла a с отсутствующим фитольным хвостом. В отличие от хлорофилла, протохлорофиллид имеет сильную флюоресценцию; мутанты, накапливающие его в своих тканях, светятся красным, если облучить их синим светом. У цветковых растений, реакция превращения протохлорофиллида в хлорофилл является светозависимой, и такие растения становятся белыми (хлорозными) если выращивать их в темноте. В отличие от них голосеменные, водоросли, и фотосинтезирующие бактерии используют другой, не зависящий от света фермент, и вырастают зелёными даже в темноте.

Светособирающие комплексы — пигмент-белковые комплексы фотосинтезирующих организмов, локализованные в фотосинтетических мембранах и выполняющие функцию первичного поглощения квантов света с последующей миграцией энергии возбуждения к реакционным центрам фотосистем. Также они обеспечивают тонкую настройку фотосинтетического аппарата и участвуют в его защите от фотоповреждений.

Фотосисте́ма I, или пластоциани́н-ферредокси́н-оксидоредукта́за — второй функциональный комплекс электрон-транспортной цепи (ЭТЦ) хлоропластов. Он принимает электрон от пластоцианина и, поглощая световую энергию, формирует сильный восстановитель П700, способный через цепь переносчиков электронов осуществить восстановление НАДФ+. Таким образом, при участии ФСI синтезируется источник электронов (НАДФН) для последующих реакций восстановления углерода в хлоропластах в цикле Кальвина. Кроме того, ФСI может осуществлять циклический транспорт электронов, сопряжённый с синтезом АТФ, обеспечивая дополнительный синтез АТФ в хлоропластах.

Фотосисте́ма II (втора́я фотосисте́ма, фотосисте́ма два, ФСII), или H2O-пластохиноноксидоредуктаза — первый функциональный комплекс электрон-транспортной цепи (ЭТЦ) хлоропластов. Он расположен в мембранах тилакоидов всех растений, водорослей и цианобактерий. Поглощая энергию света в ходе первичных фотохимических реакций, он формирует сильный окислитель — димер хлорофилла a (П680+), который через цепь окислительно-восстановительных реакций способен вызвать окисление воды.
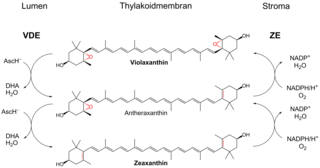
Ксантофилловый цикл, или виолаксантиновый цикл, в случае высших растений, выполняет функцию защиты фотосинтетического аппарата от избытка энергии при повышенной инсоляции. Он позволяет избежать фотоингибирования, значительно увеличив нефотохимическое тушение. Цикл включает в себя ферментативные взаимопревращения между виолаксантином и зеаксантином.

Фотоингибирование — индуцируемое светом снижение фотосинтетической активности растений, водорослей или цианобактерии. Фотосистема II (ФСII) более чувствительна к свету, чем другие фотосинтетические машины, поэтому большинство исследователей определяют термин как светоиндуцируемое повреждения ФСII. В живых организмах, поврежденные избыточным освещением фотосистемы II постоянно репарируются за счёт деградации и синтеза белка D1 фотосинтетического реакционного центра ФСII. Понятие фотоингибирование также используется в более широком смысле, в значении «динамическое фотоингибирование», чтобы описать все реакции, которые приводят к снижению эффективности фотосинтеза, когда растения подвергаются воздействию света.
Эффект Каутского — явление изменения свечения хлорофилла, которое возникает при освещении предварительно адаптированного к темноте листа растения. Этот феномен был открыт Х. Каутским и А. Хиршем в 1931 году.

Флуоресценция хлорофилла — явление свечения хлорофилла при поглощении им света, происходит в результате возвращения молекулы из возбуждённого в основное состояние. Широко используется как показатель фотосинтетического преобразования энергии у высших растений, водорослей и бактерий. Возбуждённый хлорофилл теряет поглощённую световую энергию, растрачивая её на фотосинтез, переводя её в тепло в результате нефотохимического тушения или излучая в виде флуоресценции. Поскольку все эти процессы конкурируют друг с другом, анализируя флуоресценцию хлорофилла, можно получить представления об интенсивности фотосинтеза и здоровье растения.
Acaryochloris marina (лат.) — вид симбиотических цианобактерий из порядка Synechococcales. В качестве основного пигмента фотосинтеза используют хлорофилл d, что позволяет им улавливать свет дальней красной области спектра.
BCAM, или «лютеранский антиген» — мембранный белок, гликопротеин суперсемейства иммуноглобулинов, молекула клеточной адгезии, продукт гена BCAM. Антиген группы крови Lutheran. Антиген b группы крови Auberger.
Рецептор интерлейкина 10, альфа-субъединица — белок, субъединица рецептора интерлейкина 10, продукт гена человека IL10RA.
