Хроматин
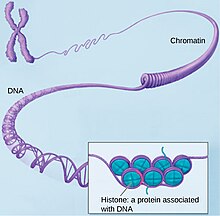
Хроматин (от греч. χρώμα «цвета; краски») — нуклеопротеид, составляющий основу хромосом. Состоит из ДНК и белков (главным образом гистонов). Хроматин находится внутри ядра клеток эукариот[1] и архей, имеющих гистоны. В более широком смысле хроматином иногда называют также и вещество нуклеоида у бактерий[2].
Именно в составе хроматина происходит реализация генетической информации, а также репликация и репарация ДНК[3].
До 25—40% сухого веса хроматина составляют гистоновые белки[1]. Гистоны являются компонентом нуклеосом, надмолекулярных структур, участвующих в упаковке хромосом. Нуклеосомы располагаются довольно регулярно, так что образующаяся структура напоминает бусы. Нуклеосома состоит из гистонов четырёх типов: H2A, H2B, H3 и H4. Эти гистоны называются ко́ровыми. В одну нуклеосому входят по два ко́ровых гистона каждого типа — всего восемь белков. Линкерный гистон H1, более крупный, чем ко́ровые гистоны, связывается с ДНК в месте её входа на нуклеосому[4].
По классическим представлениям, нить ДНК с нуклеосомами образует нерегулярную соленоид-подобную структуру толщиной около 30 нанометров, так называемую 30-нм фибриллу. В настоящее время, однако, показано, что 30-нм фибрилла является артефактом исследований in vitro, в клетках нуклеосомная фибрилла (у эукариот и некоторых архей) или непосредственно ДНК (у бактерий и некоторых архей) складывается в нерегулярные петлевые и глобулярные структуры, которые в ряде случае имеют регуляторное значение[5]. Дальнейшая упаковка может иметь различную плотность. Если хроматин упакован плотно, его называют конденсированным или гетерохроматином, он хорошо видим под микроскопом. ДНК, находящаяся в гетерохроматине не транскрибируется, обычно это состояние характерно для незначащих или молчащих участков. В интерфазе гетерохроматин обычно располагается по периферии ядра (пристеночный гетерохроматин). Полная конденсация хромосом происходит перед делением клетки.
Если хроматин упакован неплотно, его называют эу- или интерхроматином. Этот вид хроматина гораздо менее плотный при наблюдении под микроскопом и обычно характеризуется транскрипционной активностью. Плотность упаковки хроматина во многом определяется модификациями гистонов — ацетилированием, фосфорилированием, метилированием и другими модификациями.
Считается, что в ядре существуют так называемые функциональные домены хроматина (ДНК одного домена содержит приблизительно 30 тысяч пар оснований), то есть каждый участок хромосомы имеет собственную «территорию». Вопрос пространственного распределения хроматина в ядре изучен пока недостаточно. Известно, что теломерные (концевые) и центромерные (отвечающие за связывание сестринских хроматид в митозе) участки хромосом закреплены на белках ядерной ламины.
Классификация хроматина
Участки хроматина различаются по ряду взаимосвязанных параметров: уровню конденсации, активности, функциям, модификациям гистонов и др. Для систематизации широко используется деление на деконденсированный активный эухроматин и конденсированный неактивный гетерохроматин, последний подразделяют на конститутивный и факультативный. Кроме того, разработаны более детальные классификации, основывающиеся преимущественно на модификациях гистонов и рассматривающие активность и плотность хроматина как их производные. Применение в геномике нашли системы цветовой маркировки типов хроматина: 5-цветная[6] и 9-цветная[7].
Открытый хроматин
Открытый хроматин — это области ДНК, лишенные нуклеосом и гиперчувствительные к обработке ДНКазой I. Области открытого хроматина часто связаны с регуляторными участками ДНК.
Схема конденсации хроматина

Петли
Хроматин в геноме человека содержит около 10 тысяч петель, они способны исчезать и появляться снова. Предполагается, что петли способны участвовать в активации генов. Возникновению петель способствуют два белка: CTCF и когезин[8].
Примечания
- ↑ 1 2 Биологический энциклопедический словарь / Гл.ред. М.С.Гиляров. — М.: Сов. энциклопедия, 1986. — 831 с.
- ↑ Из чего же сделан хроматин архей? Дата обращения: 21 апреля 2022. Архивировано 15 мая 2022 года.
- ↑ Жимулев И. Ф. Общая и молекулярная генетика. — 1. — Новосибирск: Издательство Новосибирского университета, 2002. — 459 с. — 2000 экз. — ISBN 5761505096.
- ↑ Bruce Alberts, Alexander Johnson, Julian Lewis, Martin Raff, Keith Roberts, Peter Walter. Molecular Biology of the Cell. — 5. — Garland Science, 2008. — 1392 с. — ISBN 0815341059.
- ↑ Razin S. V., Gavrilov A. A. Chromatin without the 30-nm fiber: constrained disorder instead of hierarchical folding. (англ.) // Epigenetics. — 2014. — May (vol. 9, no. 5). — P. 653—657. — doi:10.4161/epi.28297. — PMID 24561903.
- ↑ Filion G. J., van Bemmel J. G., Braunschweig U., Talhout W., Kind J., Ward L. D., Brugman W., de Castro I. J., Kerkhoven R. M., Bussemaker H. J., van Steensel B. Systematic protein location mapping reveals five principal chromatin types in Drosophila cells. (англ.) // Cell. — 2010. — 15 October (vol. 143, no. 2). — P. 212—224. — doi:10.1016/j.cell.2010.09.009. — PMID 20888037.
- ↑ Kharchenko P. V., Alekseyenko A. A., Schwartz Y. B., Minoda A., Riddle N. C., Ernst J., Sabo P. J., Larschan E., Gorchakov A. A., Gu T., Linder-Basso D., Plachetka A., Shanower G., Tolstorukov M. Y., Luquette L. J., Xi R., Jung Y. L., Park R. W., Bishop E. P., Canfield T. K., Sandstrom R., Thurman R. E., MacAlpine D. M., Stamatoyannopoulos J. A., Kellis M., Elgin S. C., Kuroda M. I., Pirrotta V., Karpen G. H., Park P. J. Comprehensive analysis of the chromatin landscape in Drosophila melanogaster. (англ.) // Nature. — 2011. — 24 March (vol. 471, no. 7339). — P. 480—485. — doi:10.1038/nature09725. — PMID 21179089.
- ↑ Эрез Либерман Эйден. Распутать геном // В мире науки. — 2019. — № 5-6. — С. 58-66.
Литература
- Suhas S. P. Rao, et al.(2014). A 3D map of the human genome at kilobase resolution reveals principles of chromatin looping. Cell, doi:10.1016/j.cell.2014.11.021
- Jesse R. Dixon, David U. Gorkin, Bing Ren (2016). Chromatin Domains: The Unit of Chromosome Organization. Molecular Cell. doi:10.1016/j.molcel.2016.05.018 Обзор, посвящённый структурной организации доменов хроматина
- Tom Misteli (2020). The Self-Organizing Genome: Principles of Genome Architecture and Function. Cell. doi:10.1016/j.cell.2020.09.014 (К статье приложен отличный список литературы по теме)
См. также
| Ядерная мембрана/ Ядерная ламина |  | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Ядрышко | |||||||||
| Прочее |
| ||||||||
| Основное |  | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Классификация | |||||||||||
| Структура |
| ||||||||||
| Перестройки и нарушения | |||||||||||
| Хромосомное определение пола | |||||||||||
| Методы | |||||||||||