Хромосома

Хромосо́мы (др.-греч. χρῶμα «цвет» + σῶμα «тело») — нуклеопротеидные структуры в ядре эукариотической клетки, в которых сосредоточена бо́льшая часть наследственной информации и которые предназначены для её хранения, реализации и передачи. Хромосомы чётко различимы в световом микроскопе только в период митотического или мейотического деления клетки. Набор всех хромосом клетки, называемый кариотипом, является видоспецифичным признаком, для которого характерен относительно низкий уровень индивидуальной изменчивости[1].
Хромосома эукариот образуется из единственной и чрезвычайно длинной молекулы ДНК, которая содержит линейную группу множества генов. Необходимыми функциональными элементами хромосомы эукариот являются центромера, теломеры и точки инициации репликации. Точки начала репликации (сайты инициации) и теломеры, находящиеся на концах хромосом, позволяют молекуле ДНК эффективно реплицироваться, тогда как в центромерах сестринские молекулы ДНК прикрепляются к митотическому веретену деления, что обеспечивает их точное расхождение по дочерним клеткам в митозе[2].
Исходно термин был предложен для обозначения структур, выявляемых в эукариотических клетках, но в последние десятилетия всё чаще говорят о бактериальных или вирусных хромосомах. Поэтому, по мнению Д. Е. Корякова и И. Ф. Жимулёва[3], более широким определением является определение хромосомы как структуры, которая содержит нуклеиновую кислоту и функция которой состоит в хранении, реализации и передаче наследственной информации. Хромосомы эукариот — это ДНК-содержащие структуры в ядре, митохондриях и пластидах. Хромосомы прокариот — это ДНК-содержащие структуры в клетке без ядра. Хромосомы вирусов — это молекула ДНК или РНК внутри капсида.
История открытия хромосом
Первые описания хромосом появились в статьях и книгах разных авторов в 70-х годах XIX века, и приоритет открытия хромосом отдают разным людям, а именно: И. Д. Чистякову (1873), А. Шнейдеру (1873), Э. Страсбургеру (1875), О. Бючли (1876) и другим[4]. Чаще всего годом открытия хромосом называют 1882 год, а их первооткрывателем — немецкого анатома В. Флемминга, который в своей фундаментальной книге «Zellsubstanz, Kern und Zelltheilung» собрал и упорядочил сведения о хромосомах, дополнив результатами собственных исследований. Термин «хромосома» был предложен немецким гистологом Г. Вальдейером в 1888 году. «Хромосома» в буквальном переводе означает «окрашенное тело», поскольку осно́вные красители, такие как азуры, основной фуксин, орсеин и др., хорошо связываются хромосомами[5].
После переоткрытия в 1900 году законов Менделя потребовалось всего один-два года для того, чтобы стало ясно, что хромосомы при мейозе и оплодотворении ведут себя именно так, как это ожидалось от «частиц наследственности». В 1902 году Т. Бовери и в 1902—1903 годах У. Сеттон (Walter Sutton) независимо друг от друга выдвинули гипотезу о генетической роли хромосом[6].
Экспериментальное подтверждение этих идей было осуществлено в первой четверти XX века американскими учёными Т. Морганом, К. Бриджесом, А. Стёртевантом и Г. Мёллером. Объектом их генетических исследований послужила плодовая мушка D.melanogaster. На основе данных, полученных на дрозофиле, они сформулировали «хромосомную теорию наследственности», согласно которой передача наследственной информации связана с хромосомами, в которых линейно, в определённой последовательности, локализованы гены. Основные положения хромосомной теории наследственности были опубликованы в 1915 году в книге «The mechanism of mendelian heredity» (англ.)[7][6].
В 1933 году за открытие роли хромосом в наследственности Т. Морган получил Нобелевскую премию по физиологии и медицине[8].
Морфология метафазных хромосом


В ходе клеточного цикла облик хромосомы меняется. В интерфазе это очень нежные структуры, занимающие в ядре отдельные хромосомные территории, но не заметные как обособленные образования при визуальном наблюдении. В митозе хромосомы преобразуются в плотно упакованные элементы, способные сопротивляться внешним воздействиям, сохранять свою целостность и форму[9][10]. Именно хромосомы на стадии профазы, метафазы или анафазы митоза доступны для наблюдения с помощью светового микроскопа. Митотические хромосомы можно увидеть у любого организма, клетки которого способны делиться митозом, исключение составляют дрожжи S.cerevisiae, чьи хромосомы слишком малы[11]. Обычно митотические хромосомы имеют размеры в несколько микрон. Например, самая большая хромосома человека, 1-я хромосома имеет длину около 7—8 мкм в метафазе и 10 мкм в профазе митоза[12].

На стадии метафазы митоза хромосомы состоят из двух продольных копий, которые называются сестринскими хроматидами и которые образуются при репликации. У метафазных хромосом сестринские хроматиды соединены в районе первичной перетяжки, называемой центромерой. Центромера отвечает за расхождение сестринских хроматид в дочерние клетки при делении. На центромере происходит сборка кинетохора — сложной белковой структуры, определяющей прикрепление хромосомы к микротрубочкам веретена деления — движителям хромосомы в митозе[13]. Центромера делит хромосомы на две части, называемые плечами. У большинства видов короткое плечо хромосомы обозначают буквой p, длинное плечо — буквой q. Длина хромосомы и положение центромеры являются основными морфологическими признаками метафазных хромосом.
В зависимости от расположения центромеры различают три типа строения хромосом:
- акроцентрические хромосомы, у которых центромера находится практически на конце, и второе плечо настолько мало, что его может быть не видно на цитологических препаратах;
- субметацентрические хромосомы с плечами неравной длины;
- метацентрические хромосомы, у которых центромера расположена посередине или почти посередине[14].
Эту классификацию хромосом на основе соотношения длин плеч предложил в 1912 году российский ботаник и цитолог С. Г. Навашин. Помимо вышеуказанных трёх типов С. Г. Навашин выделял ещё и телоцентрические хромосомы, то есть хромосомы только с одним плечом. Однако по современным представлениям истинно телоцентрических хромосом не бывает. Второе плечо, пусть даже очень короткое и невидимое в обычный микроскоп, всегда присутствует[15].
Дополнительным морфологическим признаком некоторых хромосом является так называемая вторичная перетяжка, которая внешне отличается от первичной отсутствием заметного угла между сегментами хромосомы. Вторичные перетяжки бывают различной длины и могут располагаться в различных точках по длине хромосомы. Во вторичных перетяжках находятся, как правило, ядрышковые организаторы, содержащие многократные повторы генов, кодирующих рибосомные РНК. У человека вторичные перетяжки, содержащие рибосомные гены, находятся в коротких плечах акроцентрических хромосом, они отделяют от основного тела хромосомы небольшие хромосомные сегменты, называемые спутниками[16]. Хромосомы, обладающие спутником, принято называть SAT-хромосомами (лат. SAT (Sine Acid Thymonucleinico) — без ДНК).
Дифференциальная окраска метафазных хромосом

При монохромном окрашивании хромосом (ацетокармином, ацетоорсеином, окрашиванием по Фёльгену или Романовскому — Гимзе) можно идентифицировать число и размеры хромосом; их форму, определяемую прежде всего положением центромер, наличием вторичных перетяжек, спутников. В подавляющем числе случаев для идентификации индивидуальных хромосом в хромосомном наборе этих признаков недостаточно. Кроме того, монохромно окрашенные хромосомы часто очень похожи у представителей разных видов. Дифференциальное окрашивание хромосом, различные методики которого были разработаны в начале 70-х годов XX века, снабдило цитогенетиков мощнейшим инструментом для идентификации как индивидуальных хромосом в целом, так и их частей, облегчив тем самым процедуру анализа генома[17].
Методы дифференциального окрашивания делятся на две основные группы:
- методы селективного окрашивания определённых хромосомных районов, таких как блоки конститутивного гетерохроматина, активные ядрышкообразующие районы, центромерные и теломерные районы;
- методы дифференциального окрашивания эухроматиновых районов хромосом, обеспечивающие выявление в эухроматиновых районах чередующихся сегментов, так называемых бэндов (англ. band — полоса, лента, тесьма), которые окрашиваются с различной интенсивностью[18].
Уровни компактизации хромосомной ДНК

Основу хромосомы составляет линейная макромолекула ДНК значительной длины. В молекулах ДНК хромосом человека насчитывается от 50 до 245 миллионов пар азотистых оснований. Суммарная длина всех молекул ДНК из ядра человеческой клетки составляет величину около двух метров. При этом типичное ядро клетки человека, которое можно увидеть только при помощи микроскопа, занимает объём около 110 мкм³, а митотическая хромосома человека в среднем не превышает 5—6 мкм. Подобная компактизация генетического материала возможна благодаря наличию у эукариот высокоорганизованной системы укладки молекул ДНК как в интерфазном ядре, так и в митотической хромосоме. У эукариот в пролиферирующих клетках осуществляется постоянное закономерное изменение степени компактизации хромосом. Перед митозом хромосомная ДНК компактизуется в 105 раз по сравнению с линейной длиной ДНК, что необходимо для успешной сегрегации хромосом в дочерние клетки, в то время как в интерфазном ядре для успешного протекания процессов транскрипции и репликации хромосоме необходимо декомпактизоваться[12]. При этом ДНК в ядре никогда не бывает полностью вытянутой и всегда в той или иной степени упакована. Так, расчётное уменьшение размера между хромосомой в интерфазе и хромосомой в митозе составляет всего примерно 2 раза у дрожжей и 4—50 раз у человека[19].
Упаковка ДНК в хроматин обеспечивает многократное сокращение линейных размеров ДНК, необходимое для размещения её в ядре. По классическим представлениям, упаковка имеет иерархический характер. Наиболее изученными являются три первых уровня упаковки: (1) накручивание ДНК на нуклеосомы с образованием нуклеосомной нити диаметром 10 нм, (2) компактизация нуклеосомной нити с образованием так называемой 30-нм фибриллы и (3) сворачивание последней в гигантские (50 — 200 тысяч п. н.) петли, закреплённые на белковой скелетной структуре ядра — ядерном матриксе[20].
По современным представлениям, однако, такие регулярные структуры — артефакты, образующиеся в нефизиологических условиях in vitro. В клетках нуклеосомная фибрилла (у эукариот и некоторых архей) или непосредственно ДНК (у бактерий и некоторых архей) складывается в петлевые и глобулярные структуры, которые в ряде случае имеют регуляторное значение[21]
Одним из самых последних уровней упаковки в митотическую хромосому некоторые исследователи, придерживающиеся традиционных взглядов, считают так называемую хромонему, толщина которой составляет около 0,1—0,3 мкм[22]. В результате дальнейшей компактизации диаметр хроматиды достигает ко времени метафазы 700 нм. Значительная толщина хромосомы (диаметр 1400 нм) на стадии метафазы позволяет, наконец, увидеть её в световой микроскоп. Конденсированная хромосома имеет вид буквы X (часто с неравными плечами), поскольку две хроматиды, возникшие в результате репликации, соединены между собой в районе центромеры (подробнее о судьбе хромосом при клеточном делении см. статьи митоз и мейоз).
Хромосомные аномалии
Анеуплоидия
При анеуплоидии происходит изменение числа хромосом в кариотипе, при котором общее число хромосом не кратно гаплоидному хромосомному набору n. В случае утраты одной хромосомы из пары гомологичных хромосом мутантов называют моносомиками, в случае одной дополнительной хромосомы мутантов с тремя гомологичными хромосомами называют трисомиками, в случае утраты одной пары гомологов — нуллисомиками[23]. Анеуплоидия по аутосомным хромосомам всегда вызывает значительные нарушения развития, являясь основной причиной спонтанных абортов у человека[24]. Одной из самых известных анеуплоидий у человека является трисомия по хромосоме 21, которая приводит к развитию синдрома Дауна[1]. Анеуплоидия характерна для опухолевых клеток, особенно для клеток со́лидных опухолей[25].
Полиплоидия
Изменение числа хромосом, кратное гаплоидному набору хромосом (n), называется полиплоидией. Полиплоидия широко и неравномерно распространена в природе. Известны полиплоидные эукариотические микроорганизмы — грибы и водоросли, часто встречаются полиплоиды среди цветковых, но не среди голосеменных растений. Полиплоидия клеток всего организма у многоклеточных животных редка, хотя у них часто встречается эндополиплоидия некоторых дифференцированных тканей, например, печени у млекопитающих, а также тканей кишечника, слюнных желёз, мальпигиевых сосудов ряда насекомых[26].
Хромосомные перестройки
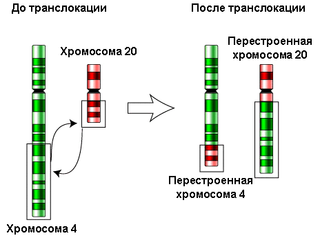
Хромосомные перестройки (хромосомные аберрации) — это мутации, нарушающие структуру хромосом. Они могут возникнуть в соматических и зародышевых клетках спонтанно или в результате внешних воздействий (ионизирующее излучение, химические мутагены, вирусная инфекция и др.). В результате хромосомной перестройки может быть утрачен или, наоборот, удвоен фрагмент хромосомы (делеция и дупликация, соответственно); участок хромосомы может быть перенесён на другую хромосому (транслокация) или он может изменить свою ориентацию в составе хромосомы на 180° (инверсия). Существуют и другие хромосомные перестройки.
Необычные типы хромосом



Микрохромосомы
У многих птиц и рептилий хромосомы в кариотипе образуют две чёткие группы: макрохромосомы и микрохромосомы. У некоторых видов микрохромосомы настолько мелкие и их так много, что невозможно отличить одну от другой[27]. Микрохромосомы являются короткими по длине, но обогащёнными генами хромосомами. Например, кариотип курицы содержит 39 пар хромосом, 6 из которых относятся к макрохромосомам, а 33 — к минихромосомам. Макрохромосомы курицы содержат две трети геномной ДНК, но только 25 % генов, в то время как микрохромосомы содержат оставшуюся треть геномной ДНК и 75 % генов. Таким образом, плотность генов в минихромосомах курицы в шесть раз выше, чем в макрохромосомах[28].
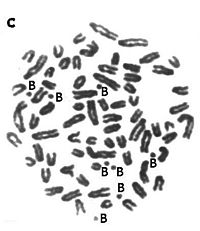
B-хромосомы
B-хромосомы — это добавочные хромосомы, которые имеются в кариотипе только у отдельных особей в популяции. Они часто встречаются у растений, описаны у грибов, насекомых и животных. Некоторые В-хромосомы содержат гены, часто это гены рРНК, однако не ясно, насколько эти гены функциональны. Наличие В-хромосом может влиять на биологические характеристики организмов, особенно у растений, где их наличие ассоциируется с пониженной жизнеспособностью. Предполагается, что В-хромосомы постепенно утрачиваются в соматических клетках в результате нерегулярности их наследования[28].
Голоцентрические хромосомы
Голоцентрические хромосомы не имеют первичной перетяжки, они имеют так называемый диффузный кинетохор, поэтому во время митоза микротрубочки веретена деления прикрепляются по всей длине хромосомы. Во время расхождения хроматид к полюсам деления у голоцентрических хромосом они идут к полюсам параллельно друг другу, в то время как у моноцентрической хромосомы кинетохор опережает остальные части хромосомы, что приводит к характерной V-образной форме расходящихся хроматид на стадии анафазы. При фрагментации хромосом, например, в результате воздействия ионизирующего излучения, фрагменты голоцентрических хромосом расходятся к полюсам упорядоченно, а не содержащие центромеры фрагменты моноцентрических хромосом распределяются между дочерними клетками случайным образом и могут быть утрачены[29].
Голоцентрические хромосомы встречаются у протист, растений и животных. Голоцентрическими хромосомами обладает нематода C. elegans[30].
Гигантские формы хромосом
Политенные хромосомы
Политенные хромосомы — это гигантские скопления объединённых хроматид, возникающие в некоторых типах специализированных клеток. Впервые описаны Эдуардом-Жераром Бальбиани (фр. Édouard-Gérard Balbiani) в 1881 году в клетках слюнных желёз мотыля (Chironomus), их исследование было продолжено уже в 1930-х годах Костовым, Пейнтером, Хайнцем (нем. Emil Heintz) и Бауэром (Hans Bauer). Политенные хромосомы обнаружены также в клетках слюнных желёз, кишечника, трахей, жирового тела и мальпигиевых сосудов личинок двукрылых.
Хромосомы типа ламповых щёток
Хромосомы типа ламповых щёток — это гигантская форма хромосом, которая возникает в мейотических женских клетках на стадии диплотены профазы I у некоторых животных, в частности, у некоторых земноводных и птиц[31]. Эти хромосомы являются крайне транскрипционно активными и наблюдаются в растущих ооцитах тогда, когда процессы синтеза РНК, приводящие к образованию желтка, наиболее интенсивны. В настоящее время известно 45 видов животных, в развивающихся ооцитах которых можно наблюдать такие хромосомы. Хромосомы типа ламповых щёток не образуются в ооцитах млекопитающих[32].
Впервые хромосомы типа ламповых щёток были описаны В. Флеммингом в 1882 году. Название «хромосомы типа ламповых щёток» было предложено немецким эмбриологом И. Рюккертом (J. Rϋckert) в 1892 году.
По длине хромосомы типа ламповых щёток превышают политенные хромосомы. Например, общая длина хромосомного набора в ооцитах некоторых хвостатых амфибий достигает 5900 мкм.
Бактериальные хромосомы
Прокариоты (археи и бактерии, в том числе митохондрии и пластиды, постоянно обитающие в клетках большинства эукариот) не имеют хромосом в собственном смысле этого слова. У большинства из них в клетке имеется только одна макромолекула ДНК, замкнутая в кольцо (эта структура получила название нуклеоид). У ряда бактерий обнаружены линейные (не замкнутые в кольцо) макромолекулы ДНК. Помимо нуклеоида или линейных макромолекул, ДНК может присутствовать в цитоплазме прокариотных клеток в виде небольших замкнутых в кольцо молекул ДНК, так называемых плазмид, содержащих обычно незначительное, по сравнению с бактериальной хромосомой, число генов. Состав плазмид может быть непостоянен, бактерии могут обмениваться плазмидами в ходе парасексуального процесса.
Известно о наличии у бактерий белков, связанных с ДНК нуклеоида, но гистонов у них не обнаружено.
Хромосомы человека
Нормальный кариотип человека представлен 46 хромосомами. Это 22 пары аутосом и одна пара половых хромосом (XY в мужском кариотипе и XX — в женском). В приведённой ниже таблице показано число генов и оснований в хромосомах человека.

| Хромосома | Всего пар оснований[34] | Количество генов[35] | Количество белок-кодирующих генов[36] |
|---|---|---|---|
| 1 | 249250621 | 3511 | 2076 |
| 2 | 243199373 | 2368 | 1329 |
| 3 | 198022430 | 1926 | 1077 |
| 4 | 191154276 | 1444 | 767 |
| 5 | 180915260 | 1633 | 896 |
| 6 | 171115067 | 2057 | 1051 |
| 7 | 159138663 | 1882 | 979 |
| 8 | 146364022 | 1315 | 702 |
| 9 | 141213431 | 1534 | 823 |
| 10 | 135534747 | 1391 | 774 |
| 11 | 135006516 | 2168 | 1914 |
| 12 | 133851895 | 1714 | 1068 |
| 13 | 115169878 | 720 | 331 |
| 14 | 107349540 | 1532 | 862 |
| 15 | 102531392 | 1249 | 615 |
| 16 | 90354753 | 1326 | 883 |
| 17 | 81195210 | 1773 | 1209 |
| 18 | 78077248 | 557 | 289 |
| 19 | 59128983 | 2066 | 1492 |
| 20 | 63025520 | 891 | 561 |
| 21 | 48129895 | 450 | 246 |
| 22 | 51304566 | 855 | 507 |
| X-хромосома | 155270560 | 1672 | 837 |
| Y-хромосома | 59373566 | 429 | 76 |
| Всего | 3 079 843 747 | 36463 | 21364 |
См. также
Примечания
- ↑ 1 2 Тарантул В. З. . Толковый биотехнологический словарь. — М.: Языки славянских культур, 2009. — 936 с. — 400 экз. — ISBN 978-5-9551-0342-6.
- ↑ Молекулярная биология клетки: в 3-х томах / Б. Альбертс, А. Джонсон, Д. Льюис и др. — М.-Ижевск: НИЦ «Регулярная и хаотическая динамика», Институт компьютерных исследований, 2013. — Т. I. — 808 с. — ISBN 978-5-4344-0112-8. — С. 309—336.
- ↑ Коряков, Жимулёв, 2009, с. 13.
- ↑ Филипченко Ю. А. . Генетика. — Л.: Типография «Печатный Двор», 1929. — 379 с. Архивировано 8 сентября 2013 года.
- ↑ Коряков, Жимулёв, 2009, с. 9.
- ↑ 1 2 Коряков, Жимулёв, 2009, с. 12.
- ↑ Morgan T. H., Sturtevant A. H., Muller H. J., Bridges C. B. . The mechanism of mendelian heredity. — New York: Henry Holt and Company, 1915. — 262 с.
- ↑ The Nobel Prize in Physiology or Medicine 1933 (англ.). // Nobel Media AB 2013. Дата обращения: 11 декабря 2013. Архивировано 21 августа 2007 года.
- ↑ Рубцов Н. Б. Хромосома человека в четырёх измерениях // Природа. — Наука, 2007. — № 8. — С. 3—10. Архивировано 2 июля 2014 года.
- ↑ Рубцов Н. Б. Организация хромосом: 70 лет спустя // Природа. — Наука, 2012. — № 10. — С. 24—31. Архивировано 1 июля 2014 года.
- ↑ Коряков, Жимулёв, 2009, с. 29.
- ↑ 1 2 Смирнов А. Ф. . Структурно-функциональная организация хромосом. — СПб.: Нестор-История, 2009. — 204 с. — ISBN 978-5-98187-486-4. Архивировано 5 марта 2014 года.
- ↑ Вершинин А. В. Центромеры и теломеры хромосом // Природа. — Наука, 2007. — № 9. — С. 21—27. Архивировано 11 января 2014 года.
- ↑ Инге-Вечтомов, 2010, с. 84—87.
- ↑ Коряков, Жимулёв, 2009, с. 30.
- ↑ Pikaard C. S. The epigenetics of nucleolar dominance (англ.) // Trends in Genetics. — Cell Press, 2000. — Vol. 16, no. 11. — P. 495—500. Архивировано 21 июня 2010 года.
- ↑ Зощук Н. В., Бадаева Е. Д., Зеленин А. В. История современного хромосомного анализа. Дифференциальное окрашивание хромосом растений // Онтогенез. — 2003. — Т. 34, № 1. — С. 5—18. — PMID 12625068.
- ↑ Рубцов Н. Б. . Методы работы с хромосомами млекопитающих: Учеб. пособие. — Новосибирск: Новосиб. гос. ун-т, 2006. — 152 с. — ISBN 5-94356-376-8.
- ↑ Коряков, Жимулёв, 2009, с. 91.
- ↑ Разин С. В. . Хроматин: упакованный геном / С. В. Разин, А. А. Быстрицкий. — М.: БИНОМ: Лаборатория знаний, 2009. — 176 с. — ISBN 978-5-9963-0087-7.
- ↑ Razin S. V., Gavrilov A. A. Chromatin without the 30-nm fiber: constrained disorder instead of hierarchical folding. (англ.) // Epigenetics. — 2014. — May (vol. 9, no. 5). — P. 653—657. — doi:10.4161/epi.28297. — PMID 24561903.
- ↑ Ченцов Ю. С., Бураков В. В. Хромонема — забытый уровень укладки хроматина в митотических хромосомах // Биологические мембраны. — 2005. — Т. 22, № 3. — С. 178—187. — ISSN 0233-4755.
- ↑ Коряков, Жимулёв, 2009, с. 45—46.
- ↑ Hassold T., Hall H., Hunt P. The origin of human aneuploidy: where we have been, where we are going // Human Molecular Genetics[англ.]. — Oxford University Press, 2007. — Vol. 16, spec. no. 2. — P. R203—R208. — doi:10.1093/hmg/ddm243. — PMID 17911163.
- ↑ Holland A. J., Cleveland D. W. Losing balance: the origin and impact of aneuploidy in cancer // EMBO Reports. — 2012. — Vol. 13, no. 6. — P. 501—514. — doi:10.1038/embor.2012.55. — PMID 22565320. Архивировано 19 марта 2017 года.
- ↑ Инге-Вечтомов, 2010, с. 401—414.
- ↑ Коряков, Жимулёв, 2009, с. 31.
- ↑ 1 2 Браун Т. А. . Геномы / Пер. с англ. = Genomes. — М.-Ижевск: Институт компьютерных исследований, 2011. — 944 с. — ISBN 978-5-4344-0002-2. Архивировано 17 июля 2019 года.
- ↑ Mandrioli M., Manicardi G. C. Unlocking holocentric chromosomes: new perspectives from comparative and functional genomics? // Current Genomics. — 2012. — Vol. 13, no. 5. — P. 343—349. — doi:10.2174/138920212801619250. — PMID 23372420.
- ↑ Dernburg A. F. Here, there, and everywhere: kinetochore function on holocentric chromosomes // The Journal of Cell Biology. — 2001. — Vol. 153, no. 6. — P. F33—F38. — PMID 11402076.
- ↑ Gall J. G. Are lampbrush chromosomes unique to meiotic cells? // Chromosome Research. — 2012. — Vol. 20, no. 8. — P. 905—909. — doi:10.1007/s10577-012-9329-5. — PMID 23263880.
- ↑ Macgregor H. So what’s so special about these things called lampbrush chromosomes? // Chromosome Research. — 2012. — Vol. 20, no. 8. — P. 903—904. — doi:10.1007/s10577-012-9330-z. — PMID 23239398. Архивировано 19 марта 2017 года.
- ↑ Bolzer, Andreas; Kreth, Gregor; Solovei, Irina; Koehler, Daniela; Saracoglu, Kaan; Fauth, Christine; Müller, Stefan; Eils, Roland; Cremer, Christoph; Speicher, Michael R.; Cremer, Thomas. Three-Dimensional Maps of All Chromosomes in Human Male Fibroblast Nuclei and Prometaphase Rosettes (англ.) // PLoS Biology : journal. — 2005. — Vol. 3, no. 5. — P. e157. — doi:10.1371/journal.pbio.0030157. — PMID 15839726. — PMC 1084335.

- ↑ Human Genome Assembly Information (англ.). // Genome Reference Concortium. Дата обращения: 18 апреля 2013. Архивировано 9 апреля 2013 года.
- ↑ Homo sapiens Genome: Statistics -- Build 37.3. // NCBI. Дата обращения: 18 апреля 2013. Архивировано 13 сентября 2017 года.
- ↑ Ensembl. Location: whole genome (англ.). // The Ensembl project. Дата обращения: 25 апреля 2013. Архивировано 28 апреля 2013 года.
Литература
- Захаров А. Ф., Бенюш В. А., Кулешов Н. П., Барановская Л. И. Хромосомы человека. Атлас. — М.: Медицина, 1982. — 263 с.
- Инге-Вечтомов С. Г. Генетика с основами селекции: учебник для студентов высших учебных заведений. — СПб.: Изд-во Н-Л, 2010. — 720 с. — ISBN 978-5-94869-105-3. — С. 193—194.
- Коряков Д. Е., Жимулев И. Ф. Хромосомы. Структура и функции. — Новосибирск: Изд-во СО РАН, 2009. — 258 с. — ISBN 978-5-7692-1045-7.
- Лима-де-Фариа А. Похвала «глупости» хромосомы. — М.: БИНОМ. Лаборатория знаний, 2012. — 312 с. — ISBN 978-5-9963-0148-5.
- Молекулярная биология клетки: в 3-х томах / Б. Альбертс, А. Джонсон, Д. Льюис и др. — М.-Ижевск: НИЦ «Регулярная и хаотическая динамика», Институт компьютерных исследований, 2013. — Т. I. — 808 с. — ISBN 978-5-4344-0112-8. — С. 325—359.
| Словари и энциклопедии | ||||
|---|---|---|---|---|
| Основное |  | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Классификация | |||||||||||
| Структура |
| ||||||||||
| Перестройки и нарушения | |||||||||||
| Хромосомное определение пола | |||||||||||
| Методы | |||||||||||
| Ключевые понятия |  | |
|---|---|---|
| Области генетики | ||
| Закономерности | ||
| Связанные темы | ||