Хромосомная теория наследственности


Хромосомная теория наследственности — теория, согласно которой передача наследственной информации в ряду поколений связана с передачей хромосом, в которых в определённой и линейной последовательности расположены гены. Эта теория сформулирована в начале XX века, основной вклад в её создание внесли американский цитолог У. Саттон[англ.], немецкий эмбриолог Т. Бовери и американский генетик Т. Морган со своими сотрудниками К. Бриджесом, А. Стёртевантом и Г. Мёллером[1].
В 1902-1903 годах У. Саттон и Т. Бовери независимо друг от друга выявили параллелизм в поведении менделевских факторов наследственности (генов) и хромосом. Эти наблюдения послужили основой для предположения, что гены расположены в хромосомах. Экспериментальное доказательство локализации генов в хромосомах было получено позднее Т. Морганом и его сотрудниками, работавшими с плодовой мушкой Drosophila melanogaster[2]. Начиная с 1911 года, эта группа опытным путём доказала, что гены располагаются в хромосомах линейно; что находящиеся на одной хромосоме гены наследуются сцепленно; что сцепленное наследование может нарушаться за счёт кроссинговера[3]. Основные выводы сформулированной ими хромосомной теории наследственности были опубликованы в 1915 году в книге «Механизм менделевской наследственности»[4].
В 1933 году Томасу Моргану за открытие роли хромосом в наследственности была присуждена Нобелевская премия по физиологии и медицине[5].
По мнению Н. В. Тимофеева-Ресовского, высказанному им в 1964 году, «…вся экспериментальная генетика XX века была ничем иным, как развитием и анализом деталей хромосомной теории наследственности»[6].
Первые описания хромосом
Начальным этапом создания хромосомной теории наследственности можно считать первые описания хромосом во время деления соматических клеток, сделанных во второй половине XIX века в работах И. Д. Чистякова (1873), А. Шнейдера (1873), а главным образом Э. Страсбургера (1875) и О. Бючли (1876). Термина «хромосома» тогда ещё не существовало, и вместо него говорили о «сегментах», на которые распадается хроматиновый клубок, или о «хроматиновых элементах». Термин «хромосома» был предложен позднее Г. Вальдейером в его сводной статье 1888 года[7].
Параллельно с изучением соматических митозов шло и изучение процесса оплодотворения, как в животном, так и в растительном царстве. Слияние семенного ядра с яйцевым впервые наблюдал у иглокожих О. Гертвиг (1876), а среди растений у лилейных Страсбургер (1884). Именно на основании этих наблюдений в 1884 году оба они пришли к выводу, что клеточное ядро является носителем наследственных свойств организма[7].
Центр внимания с ядра, как целого, на его отдельные хромосомы был перенесён лишь после того, как появилась чрезвычайно важная для того времени работа Э. ван Бенедена (1883)[8]. Ему при изучении процесса оплодотворения у аскариды, имеющей очень малое число хромосом — всего 4 в соматических клетках, удалось подметить, что хромосомы в первом делении оплодотворённого яйца происходят наполовину из ядра сперматозоида и наполовину — из ядра яйцеклетки[9]. Таким образом, во-первых, был открыт факт, что половые клетки имеют вдвое меньшее количество хромосом по сравнению с соматическими клетками, а во-вторых, был впервые поставлен вопрос о хромосомах, как особых постоянных сущностях в клетке[7].
Индивидуальность хромосом. Работы Бовери
Следующий этап связан с развитием концепции индивидуальности хромосом. Одним из первых шагов было установление того, что соматические клетки разных тканей одного и того же организма обладают одинаковым числом хромосом. Первым на это ясно и вполне определённо указал австрийский зоолог Карл Рабль в 1885 году в своей статье «О клеточном делении»[10][11]. Экспериментальное обоснование закона индивидуальности хромосом было обеспечено работами немецкого зоолога Теодора Бовери, а именно — целым рядом классических исследований, посвящённых хромосомам: «Этюды о клетке» (1887—1890 и далее), «Данные о строении хроматинового вещества ядра» (1904) и др.[12][13]. Уже с самых первых работ, вышедших в 1887 и 1888 годах, Бовери решительно высказался в пользу того, что «хроматиновые элементы являются самостоятельными элементами, сохраняющими эту самостоятельность и в покоящемся ядре». Таким образом, согласно этому взгляду, хотя хромосомы ясно видны лишь во время митоза, но и на стадии интерфазы хромосомы не исчезают, а сохраняют свою обособленность и самостоятельность. Бовери также выдвинул гипотезу о качественном различии хромосом, согласно которой каждая хромосома отличается по своему внутреннему наследственному составу от других хромосом, входящих в состав того же ядра[7]. В серии блестящих экспериментов (1902—1907) над яйцами морских ежей, оплодотворённых двумя спермиями, Бовери продемонстрировал, что нарушения развития происходят при нарушении распределения хромосом. На большом статистическом материале Бовери показал, что для нормального развития требуется наличие всех хромосом, присущих виду[14].
Исследования мейоза. Гипотеза Саттона. Теория хиазмотипии
Необходимым этапом в формировании хромосомной теории наследственности было создание ясной картины преобразований хромосом в мейозе. Такая картина сложилась усилиями многих исследователей в конце XIX и в первом десятилетии XX века. Тогда же была создана и вся терминология, описывающая процесс мейоза, которой пользуются до сих пор[15].
Открывателем мейоза является упомянутый выше бельгийский исследователь Эдуард ван Бенеден, который, изучая процесс оплодотворения у аскариды лошади, в 1883 году детально описал два последовательных деления при созревании яйцеклетки и сопутствующем формировании полярных телец[9]. Благодаря малому числу хромосом у аскариды, диплоидный набор которой составляет 4 хромосомы, Э. ван Бенеден смог показать также, что у аскариды ядра гамет при оплодотворении вносят в зиготу по равному и гаплоидному набору хромосом. Феномен мейоза с его основным следствием — редукцией числа хромосом — был предсказан А. Вейсманом вскоре (1887) после этого открытия Э. ван Бенедена[15].
В 1901 году Томас Монтгомери (Thomas H. Montgomery), изучавший мейоз у онихофор Peripatus и некоторых насекомых, пришёл к важному заключению, что конъюгирующие в профазе I мейоза хромосомы представляют собой пару гомологичных хромосом материнского и отцовского происхождения[16][17].
Спустя два года после переоткрытия законов Менделя, в 1902 году, исследования процесса сперматогенеза у кузнечика кобылки Brachystola magna привели молодого американского зоолога Уолтера Саттона[англ.] к смелой гипотезе, что распределение по дочерним клеткам хромосом при гаметогенезе может представлять основу менделевского закона независимого наследования признаков[18]. В своей работе он писал:
 | Я хочу привлечь внимание к вероятности, что соединение пары отцовских и материнских хромосом и их последующее разделение во время редукционного деления может представлять физическую основу закона наследственности Менделя [14] |
Однако предположение Саттона не встретило немедленного признания, прежде всего в силу спекулятивности его характера. Также считалось загадкой, как выйти из противоречия, связанного с ограниченностью числа пар хромосом и чрезвычайно большого числа признаков[20].
В 1909 году бельгийский цитолог Франс Янсенс (Frans Alfons Janssens) на основании своих наблюдений над первым делением мейоза у калифорнийской саламандры Batrachoseps attenuatus предложил теорию взаимообмена хромосом, названной им теорией хиазмотипии. Согласно этой теории, в профазе I мейоза бок о бок конъюгируют гомологичные хромосомы материнского и отцовского происхождения, каждая из которых состоит из двух хроматид. При этом хромосомы перекручиваются вокруг друг друга, рвутся и вновь соединяются в точке перекрёста (хиазмы) таким образом, что в двух хроматидах из четырёх реципрокно объединяются сегменты материнского и отцовского происхождения. Хиазмотипия, по мнению Янсенса, могла служить цитологическим основанием менделевского закона независимого наследования признаков[21][22].
Открытие сцепленного наследования
В своей статье 1903 года «Хромосомы в наследственности» У. Саттон предположил, что одна хромосома должна содержать несколько генов, называемых им аллеломорфами, которые должны наследоваться совместно[23]. Такой вид наследования — сцепленное наследование — открыли в 1905 году Уильям Бэтсон с учениками, назвав его «гаметическое сцепление» (англ. gametic coupling). В экспериментах с душистым горошком Lathyrus odoratus они, изучая наследование цвета лепестков и формы пыльцы, обнаружили, что для этой пары признаков не наблюдается независимого наследования, то есть пара признаков, характеризующее родительское растение, имеет тенденцию наследоваться совместно, однако эти признаки не являются полностью сцепленными[24]. Для объяснения обнаруженного им явления Бэтсон создал крайне искусственную теорию редупликации, не приняв во внимание гипотезу, высказанную молодым У. Саттоном[20]. Связь между хромосомами и явлением гаметического сцепления предположил в 1906 году английский ботаник Роберт Локк (Robert Heath Lock) в своей книге «Recent Progress in the Study of Variation, Heredity, and Evolution», которая послужила учебником генетики для многих учёных того времени, включая таких известных генетиков, как Герман Мёллер и Роберт Фишер[25][4].
Открытие половых хромосом
Важным источником доказательства роли хромосом в наследственности явилось обнаружение особых, «половых» хромосом, имеющих отношение к определению пола. К началу XX века несколькими исследователями были описаны «добавочные» хромосомы у насекомых. Американский исследователь Кларенс Мак-Кланг (C. E. McClung) в 1904 году высказал догадку, что эти хромосомы, наблюдавшиеся только у половины спермиев у самцов разных видов насекомых, являются «определителями пола». В 1905 году подтверждение этой догадки почти одновременно получили Эдмунд Вильсон, обнаруживший у клопа Protenor различие в числе хромосом у самцов и самок (хромосомное определение пола, тип XO/XX), и Нетти Стивенс, выявившая различия по одной паре хромосом между самцами и самками у жука Tenebrio (хромосомное определение пола, тип XY/XX)[14]. В своих работах Мак-Кланг, Вильсон и Стивенс установили наличие полной корреляции между поведением в мейозе половых хромосом и определением пола[15].
Работы Моргана и его школы
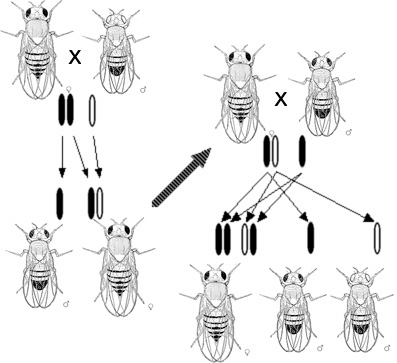
В 1909 году в лаборатории американского зоолога Томаса Ханта Моргана в Колумбийском университете начали использовать для генетических экспериментов плодовую мушку Drosophila melanogaster[26]. Многочисленные мутации, проявившиеся при лабораторном разведении дрозофилы, позволили в первую очередь обнаружить гены, наследовавшиеся «сцепленно с полом». Первой описанной мутацией стала мутация w (от англ. white — белый), обуславливающая белый цвет глаз у мушки. Публикация об этой мутации появилась в 1910 году, в ней Морган отметил, что характер наследования мутации w совпадает с наследованием хромосом, определяющих пол у дрозофилы[27][28]. Вскоре было описано ещё две сцепленных с полом мутации, и при изучении их совместного наследования Морган пришёл к заключению, что гены должны быть организованы на хромосоме линейно, и их сцепленное наследование может нарушаться из-за кроссинговера, происходящего так, как предположил ранее в своей теории хиазмотипии Янсенс[29]. В 1913 году Альфред Стёртевант, ученик Моргана, расположил шесть сцепленных с полом генов на первой генетической карте в порядке, соответствующем тому, насколько часто мутации этих генов наследуются совместно[30]. Одно из самых элегантных доказательств связи между генами и хромосомами получил другой ученик Моргана, Кэлвин Бриджес. Он соотнёс редкие случаи исключений при наследовании мутаций, сцепленных с полом, с неправильным расхождением Х-хромосом при мейозе у самок дрозофилы. Он описал самок дрозофилы c аномальным кариотипом XXY вместо нормального XX, при этом по сцепленным с полом признакам они являлись полными копиями своих матерей, что говорило о том, что обе Х-хромосомы были унаследованы от матери. Тем самым эксперименты по скрещиванию были подкреплены цитологическими наблюдениями[31].
Основные положения хромосомной теории наследственности
Анализ явлений сцепленного наследования, кроссинговера, сравнение генетической и цитологической карт позволяют сформулировать основные положения хромосомной теории наследственности:
- Гены находятся в хромосомах.
- Гены расположены в хромосоме в линейной последовательности.
- Различные хромосомы содержат неодинаковое число генов. Кроме того, набор генов каждой из негомологичных хромосом уникален.
- Аллели генов занимают одинаковые локусы в гомологичных хромосомах.
- Гены одной хромосомы образуют группу сцепления, то есть наследуются преимущественно сцеплено (совместно), благодаря чему происходит сцепленное наследование некоторых признаков. Число групп сцепления равно гаплоидному числу хромосом данного вида (у гомогаметного пола) или больше на 1 (у гетерогаметного пола).
- Сцепление нарушается в результате кроссинговера, частота которого прямо пропорциональна расстоянию между генами в хромосоме (поэтому сила сцепления находится в обратной зависимости от расстояния между генами).
- Каждый биологический вид характеризуется определённым набором хромосом — кариотипом.
См. также
Примечания
- ↑ Коряков Д.Е., Жимулев И.Ф. Хромосомы. Структура и функции. — Новосибирск: Из-во СО РАН, 2009. — С. 12. — 258 с. — ISBN 978-5-7692-1045-7.
- ↑ Инге-Вечтомов, 2010, с. 153—181.
- ↑ Гайсинович, 1988, с. 240.
- ↑ 1 2 Morgan T.H., Sturtevant A.H., Muller H.J., Bridges C.B. The mechanism of mendelian heredity. — New York: Henry Holt and Company, 1915. — 262 с.
- ↑ The Nobel Prize in Physiology or Medicine 1933 (англ.). Nobel Media AB 2013. Дата обращения: 11 декабря 2013. Архивировано 21 августа 2007 года.
- ↑ Тимофеев-Ресовский Н. В. Генетика, эволюция,значение методологии в естествознании. — Екатеринбург: Токмас-Пресс, 2009. — 144 с. Архивировано 17 декабря 2013 года.
- ↑ 1 2 3 4 Филипченко Ю.А. Генетика. — Л.: Типография "Печатный Двор", 1929. — 379 с. Архивировано 8 сентября 2013 года.
- ↑ Beneden, van E. Recherches sur la maturation de l'oeuf, la fecondation et la division cellulaire.. — Leipzig, Paris, 1883.
- ↑ 1 2 Hamoir G. The discovery of meiosis by E. Van Beneden, a breakthrough in the morphological phase of heredity // Int. J. Dev. Biol. 36: 9 - 15 (1992. — 1992. — Т. 36. — С. 9—15. Архивировано 2 февраля 2014 года.
- ↑ Вермель Е. М. История учения о клетке. — М.: Наука, 1970. — 237 с.
- ↑ Carl Rabl. Über Zelltheilung (нем.) // Morphologisches Jahrbuch. — Leipzig: Verlag von Wilhelm Engelmann, 1885. — Nr. 10. — S. 214—330. Архивировано 23 сентября 2015 года.
- ↑ Boveri, T. Zellenstudien II. Die Befruchtung und Teilung des Eies von Ascaris megalocephala. — Jena Z. Naturwiss.. — 1888. — Vol. 22. — P. 685–882.
- ↑ Boveri, T. Ergebnisse über die Konstitution der chromatischen Substanz des Zellkerns. — Jena: Verlag von Gustav Fischer, 1904.
- ↑ 1 2 3 Гайсинович, 1988, с. 238.
- ↑ 1 2 3 Цитология и генетика мейоза / В. В. Хвостова, Ю. Ф. Богданов. — М.: Наука, 1975. — С. 15—27. — 432 с.
- ↑ Montgomery T.H. Some Observations and Considerations upon the Maturation Phenomena of the Germ Cells // Biological Bulletin. — 1904. — Т. 6, № 3. — С. 137—158.
- ↑ Sturtevant, 2001, p. 33—38.
- ↑ Crow E.W., Crow J.F. 100 years ago: Walter Sutton and the chromosome theory of heredity // Genetics. — 2002. — Т. 160, № 1. — С. 1—4. Архивировано 21 июня 2022 года.
- ↑ Sutton W. On the morphology of the chromosome group in Brachystola magna // Biol Bull. — 1902. — Т. 4. — С. 24—39. Архивировано 23 сентября 2015 года.
- ↑ 1 2 Гайсинович, 1988, с. 239.
- ↑ Koszul R, Meselson M, Van Doninck K, Vandenhaute J, Zickler D. The centenary of Janssens's chiasmatype theory // Genetics. — 2012. — Т. 191, № 2. — С. 309—17. Архивировано 24 сентября 2015 года.
- ↑ Janssens F. A. , Koszul R., Zickler D. La Theorie de la Chiasmatypie: Nouvelle interprétation des cinèses de maturation // Genetics. — 2012. — Т. 191, № 2. — С. 319—346.
- ↑ Sutton W.S. The chromosomes in heredity (англ.) // The Biological Bulletin. — 1903. — Vol. 4, no. 5. — P. 231—250. Архивировано 23 сентября 2015 года.
- ↑ McPeek, M.S. An introduction to recombination and linkage analysis // Genetic Mapping and DNA Sequencing / Terry Speed, Michael S. Waterman. — Springer, 1996. — P. 1—14. — ISBN 978-1-4612-6890-1.
- ↑ Edwards A.W. Robert Heath Lock and his textbook of genetics, 1906. // Genetics. — 2013. — Т. 194, № 3. — С. 529—37. — doi:10.1534/genetics.113.151266. Архивировано 24 сентября 2015 года.
- ↑ Thomas H. Morgan - Biographical (англ.). Nobelprize.org. Nobel Media AB 2013. Дата обращения: 1 апреля 2014. Архивировано 7 апреля 2014 года.
- ↑ Miko I. Thomas Hunt Morgan and Sex Linkage (англ.) // Nature Education. — 2008. — Vol. 1, no. 1. — P. 143. Архивировано 30 мая 2014 года.
- ↑ Morgan T. H. Sex limited inheritance in Drosophila (англ.) // Science. — 1910. — Vol. 32, no. 812. — P. 120—122. Архивировано 24 сентября 2015 года.
- ↑ Morgan T. H. Random segregation versus coupling in Mendelian inheritance. — 1911. — Т. 34, № 873. — С. 384—384. Архивировано 24 сентября 2015 года.
- ↑ Sturtevant A. H. The linear arrangement of six sex‐linked factors in Drosophila, as shown by their mode of association // Journal of experimental zoology. — 1913. — Т. 14, № 1. — С. 43—59. Архивировано 21 апреля 2014 года.
- ↑ Bridges C. B. Direct proof through non-disjunction that the sex-linked genes of Drosophila are borne by the X-chromosome // Science. — 1914. — Т. 40, № 1020. — С. 107—109. Архивировано 27 июля 2014 года.
Литература
- Гайсинович А. Е. . Зарождение и развитие генетики. — М.: Наука, 1988. — 424 с. — ISBN 5-02-005265-5.
- Инге-Вечтомов С. Г. . Генетика с основами селекции: учебник для студентов высших учебных заведений. — СПб.: Изд-во Н-Л, 2010. — 720 с. — ISBN 978-5-94869-105-3. — С. 153—181.
- Sturtevant A. H. . A History of Genetics. — New York: CSHL Press, 2001. — 174 p. — ISBN 978-087969607-8.
| Ключевые понятия |  | |
|---|---|---|
| Области генетики | ||
| Закономерности | ||
| Связанные темы | ||