Центральная догма молекулярной биологии
Центральная догма молекулярной биологии — обобщающее наблюдаемое в природе правило реализации генетической информации: информация передаётся от нуклеиновых кислот к белку, но не в обратном направлении. Правило было сформулировано Френсисом Криком в 1958 году[1] и приведено в соответствие с накопившимися к тому времени данными в 1970 году[2]. Переход генетической информации последовательно от ДНК к РНК и затем от РНК к белку является универсальным для всех без исключения клеточных организмов, лежит в основе биосинтеза макромолекул. Репликации генома соответствует информационный переход ДНК → ДНК. В природе встречаются также переходы РНК → РНК и РНК → ДНК (например, у некоторых вирусов), а также изменение конформации белков, передаваемое от молекулы к молекуле.
Информация, содержащаяся в биологических последовательностях
Биополимеры — это синтезируемые живыми существами биологические полимеры. ДНК, РНК и белки относятся к линейным полимерам, которые собираются путём последовательного присоединения друг к другу отдельных элементов — мономеров. Последовательность мономеров кодирует информацию, правила передачи которой описываются центральной догмой. Информация передаётся с высокой точностью, детерминистически и один биополимер используется как шаблон для сборки другого полимера с последовательностью, которая полностью определяется последовательностью первого полимера.
Универсальные способы передачи биологической информации
В живых организмах встречаются три вида гетерогенных, то есть состоящих из разных мономеров полимера — ДНК, РНК и белок. Передача информации между ними может осуществляться девятью (3 × 3 = 9) способами. Центральная догма разделяет эти девять типов передачи информации на три группы:
- общие типы — встречающиеся у большинства живых организмов;
- специальные типы — встречающиеся в виде исключения, у вирусов, у мобильных элементов генома или в условиях биологического эксперимента;
- неизвестные типы — не обнаруженные.
| Общие | Специальные | Неизвестные |
|---|---|---|
| ДНК → ДНК | РНК → ДНК | белок → ДНК |
| ДНК → РНК | РНК → РНК | белок → РНК |
| РНК → белок | ДНК → белок | белок → белок |
Общие способы передачи информации
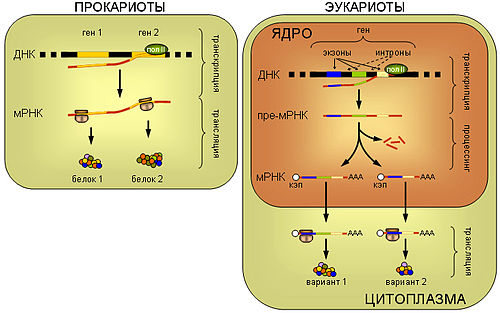
Прокариоты. У прокариот синтез белка рибосомой (трансляция) пространственно не отделён от транскрипции и может происходить ещё до завершения синтеза мРНК РНК-полимеразой. Прокариотические мРНК часто полицистронные, то есть содержат несколько независимых генов.
Эукариоты. мРНК эукариот синтезируется в виде предшественника, пре-мРНК, претерпевающего затем сложное стадийное созревание — процессинг, включающий присоединение кэп-структуры к 5'-концу молекулы, присоединение нескольких десятков остатков аденина к её 3'-концу (полиаденилирование), выщепление незначащих участков — интронов и соединение друг с другом значащих участков — экзонов (сплайсинг). При этом соединение экзонов одной и той же пре-мРНК может проходить разными способами, приводя к образованию разных зрелых мРНК, и в конечном итоге разных вариантов белка (альтернативный сплайсинг). Только мРНК, успешно прошедшая процессинг, экспортируется из ядра в цитоплазму и вовлекается в трансляцию.
Репликация ДНК: ДНК → ДНК
Репликация ДНК — основной способ передачи информации между поколениями живых организмов, поэтому точное удвоение (репликация) ДНК очень важно. Репликация осуществляется комплексом белков. Сначала белки расплетают хроматин, затем — двойную спираль; получаются две цепочки. После этого ДНК-полимераза и ассоциированные с ней белки строят на каждой из двух цепочек комплементарную ей цепочку.
Транскрипция: ДНК → РНК
Транскрипция — биологический процесс, в результате которого информация, содержащаяся на участке ДНК, копируется на синтезируемую молекулу мРНК. Транскрипцию осуществляют факторы транскрипции и РНК-полимераза. В эукариотической клетке первичный транскрипт (пре-мРНК) часто редактируется. Этот процесс называется процессингом РНК.
Трансляция: РНК → белок
Зрелая мРНК считывается рибосомами в процессе трансляции. В прокариотических клетках процесс транскрипции и трансляции не разделён пространственно, и эти процессы сопряжены. В эукариотических клетках место транскрипции — клеточное ядро — отделено от места трансляции (цитоплазмы) ядерной мембраной, поэтому мРНК транспортируется из ядра в цитоплазму. мРНК считывается рибосомой в виде трёхнуклеотидных «слов». Комплексы факторов инициации и факторов элонгации доставляют аминоацилированные транспортные РНК к комплексу мРНК-рибосома.
Специальные способы передачи информации
Обратная транскрипция: РНК → ДНК
Обратная транскрипция — перенос информации с РНК на ДНК, процесс, обратный транскрипции и осуществляемый ферментом, называемым обратной транскриптазой. Встречается у ретровирусов, например, у ВИЧ и в случае ретротранспозонов.
Репликация РНК: РНК → РНК
Репликация РНК — копирование цепи РНК на комплементарную ей цепь РНК с помощью фермента РНК-зависимой РНК-полимеразы. Таким способом реплицируются вирусы, содержащие одноцепочечную (например, пикорнавирусы, к которым относится вирус ящура, коронавирусы) или двуцепочечную РНК.
Прямая трансляция белка на матрице ДНК: ДНК → белок
Прямая трансляция была продемонстрирована в клеточных экстрактах кишечной палочки. Экстракты содержали рибосомы, но не иРНК, синтезировали белки с введённых в систему одноцепочечных ДНК; антибиотик неомицин усиливал этот эффект[3][4].
Эпигенетические изменения
Эпигенетические изменения — это изменения в проявлении генов, не обусловленные изменением генетической информации (мутациями). Эпигенетические изменения происходят в результате модификации уровня экспрессии генов, то есть их транскрипции и/или трансляции. Наиболее изученным видом эпигенетической регуляции является метилирование ДНК с помощью белков ДНК-метилтрансфераз, что приводит к временной, зависящей от условий жизни организма инактивации метилированного гена. Однако поскольку первичная структура молекулы ДНК при этом не изменяется, это исключение нельзя считать истинным примером передачи информации от белка к ДНК.
Прионы

Прионы — белки, которые существуют в двух формах. Одна из форм (конформаций) белка является функциональной, обычно растворимой в воде. Вторая форма образует нерастворимые в воде агрегаты, часто в виде молекулярных трубочек-полимеров. Мономер — молекула белка — в этой конформации способен присоединяться к другим сходным молекулам белка, переводя их во вторую, прионоподобную, конформацию. Такие процессы иногда, в отличие от всех остальных матричных процессов (первого рода), относят к матричным процессам второго рода. У грибов такие молекулы могут передаваться по наследству. Но, как и в случае метилирования ДНК, первичная структура белка в данном случае остаётся прежней, и переноса информации на нуклеиновые кислоты не происходит.
История возникновения термина «догма»
Хорас Джадсон (англ. Horace Judson) написал в книге «Восьмой день творения»:
«Я считал, что догма — это идея, которая не подтверждена фактами. Понимаете?». И Крик воскликнул с удовольствием: «Я просто не знал, что означает „догма“. Я мог бы с тем же успехом назвать это „Центральной гипотезой“ или — чем-нибудь ещё. Догма была просто слоганом».[6]
Оригинальный текст (англ.)My mind was, that a dogma was an idea for which there was no reasonable evidence. You see?!" And Crick gave a roar of delight. "I just didn't know what dogma meant. And I could just as well have called it the 'Central Hypothesis,' or — you know. Which is what I meant to say. Dogma was just a catch phrase
Кроме того, в автобиографической книге «Что за сумасшедший поиск» («What Mad Pursuit») Крик писал о выборе слова «догма» и о вызванных этим выбором проблемах:
«Я назвал эту идею центральной догмой, я подозреваю, по двум причинам. Я уже использовал слово „гипотеза“ в гипотезе о последовательности, кроме того, я хотел предположить, что это новое допущение более центральное и сильное… Как оказалось, использование термина „догма“ вызвало больше неприятностей, чем оно того стоило… Через много лет Жак Моно сказал мне, что по-видимому я не понимал, что подразумевается под словом „догма“, которая означает часть веры, не подлежащая сомнению. Я смутно опасался подобного значения слова, но поскольку я считал, что все религиозные убеждения не имеют основания, я использовал слово так, как понимал его я, а не большинство других людей, применив его к грандиозной гипотезе, которая, несмотря на внушаемое ею доверие, была основана на небольшом количестве прямых экспериментальных данных».
Оригинальный текст (англ.)I called this idea the central dogma, for two reasons, I suspect. I had already used the obvious word hypothesis in the sequence hypothesis, and in addition I wanted to suggest that this new assumption was more central and more powerful. ... As it turned out, the use of the word dogma caused almost more trouble than it was worth.... Many years later Jacques Monod pointed out to me that I did not appear to understand the correct use of the word dogma, which is a belief that cannot be doubted. I did apprehend this in a vague sort of way but since I thought that all religious beliefs were without foundation, I used the word the way I myself thought about it, not as most of the world does, and simply applied it to a grand hypothesis that, however plausible, had little direct experimental support.
См. также
- Генетический код
- ДНК
- Транскрипция
- Трансляция
- Репликация
Примечания
- ↑ Crick, F.H.C. (1958): On Protein Synthesis. Архивная копия от 24 января 2018 на Wayback Machine Symp. Soc. Exp. Biol. XII, 139—163. (pdf, early draft of original article)
- ↑ Crick, F. (1970): Central Dogma of Molecular Biology. Архивная копия от 26 января 2020 на Wayback Machine Nature 227, 561—563. PMID 4913914
- ↑ B. J. McCarthy and J. J. Holland. Denatured DNA as a Direct Template for in vitro Protein Synthesis (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 1965. — 15 September (vol. 54). — P. 880—886. — doi:10.1073/pnas.54.3.880. — PMID 4955657.
- ↑ .T. Uzawa, A. Yamagishi, T. Oshima. Polypeptide Synthesis Directed by DNA as a Messenger in Cell-Free Polypeptide Synthesis by Extreme Thermophiles, Thermus thermophilus HB27 and Sulfolobus tokodaii Strain 7 (англ.) // The Journal of Biochemistry[англ.] : journal. — 2002. — 9 April (vol. 131). — P. 849—853. — PMID 12038981.
- ↑ Андрейчук Ю. В. и др. Связь матричных процессов I и II рода: амилоиды и стабильность генома //Молекулярная биология. – 2020. – Т. 54. – №. 5. – С. 750-775.
- ↑ Horace Freeland Judson. The Eighth Day of Creation: Makers of the Revolution in Biology (25th anniversary edition) (англ.). — 1996. — ISBN 0-87969-477-7.
Ссылки
- B. J. McCarthy, J. J. Holland. Denatured DNA as a Direct Template for in vitro Protein Synthesis (англ.) // Proceedings of the National Academy of Sciences. — National Academy of Sciences, 1965. — Vol. 54. — P. 880—886.
- Werner, E. Genome Semantics, In Silico Multicellular Systems and the Central Dogma // FEBS Letters. — 2005. — Вып. 579. — С. 1779—1782. PMID 15763551
- Horace Freeland Judson. Chapter 6: My mind was, that a dogma was an idea for which there was no reasonable evidence. You see?! // The Eighth Day of Creation: Makers of the Revolution in Biology (25th anniversary edition). — 1996.