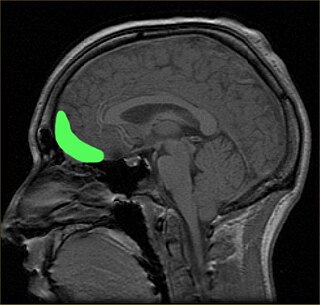
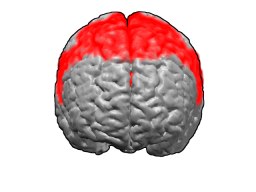
Цитоархитектоническое поле Бродмана 10
| Передняя префронтальная кора | |
|---|---|
 | |
| | |
| Часть | Лобная доля |
| Каталоги | |
Цитоархитектоническое поле Бродмана 10 (фронтополярная кора, передняя префронтальная кора) — самая передняя часть префронтальной коры в человеческом мозге[1]. 10 поле Бродмана (ПБ) — самое большое цитоархитектоническое ПБ в мозге человека. Оно также было описано как одна из наименее исследованных и понятых зон человеческого мозга[2]. Современные исследования предполагают участие 10 ПБ в стратегических процессах, в вызове необходимых воспоминаний и многих управляющих функциях. В течение человеческой эволюции функции данной зоны результировали в её увеличении относительно остального мозга[3].
Размер
Объём 10 ПБ у человека равняется в среднем 14 см³, что представляет собой 1,2 % от общего объема мозга. Такой показатель дважды превосходит стандартные ожидания касательно размера данной зоны у гоминид с объёмом мозга, свойственным Homo Sapiens. Для сравнения, объём 10 ПБ у бонобо — около 2,8 см³, что представляет собой 0,74 % от общего объёма мозга бонобо[3]. У шимпанзе обыкновенного — 2,24 см³, что является 0,57 % от общего объема мозга; у горилл — 1,94 см³, или 0,55 % от общего объема мозга; у орангутанов — 1,6 см³, или 0,45 %; у гиббонов — 0,2 см³, или 0,23 %[3].
У человека в каждом полушарии 10 ПБ содержит около 250 миллионов нейронов[3].
Фронтополярная кора человека имеет наименьшую плотность нейронов среди всех приматов[3]. Однако дендриты её нейронов крайне арборизированы, то есть очень ветвисты, и имеют очень высокую плотность дендритных шипиков[4]. Такие особенности строения являются основой для интеграции входящей информации от множества мозговых зон[2].
Связи и взаимодействия
Исследования приматов показывают, что 10 ПБ имеет обоюдные связи с другими высокоорганизованными ассоциативными зонами коры больших полушарий, в особенности, с другими зонами префронтальной коры, и в то же время 10 ПБ имеет мало связей с первичными сенсорными и моторными зонами. Нервные пути, идущие через внешнюю капсулу, связывают 10 ПБ со слуховой и мультисенсорными зонами верхней височной борозды. Другая зона, связь с которой осуществляется через внешнюю капсулу, — вентральный регион островка, или инсулярной коры. Через поясной пучок нервных волокон 10 ПБ связана с передней и задней поясной корой и ретросплениальной корой (находится сразу позади валика («splenium») мозолистого тела и включает в себя 29 и 30 ПБ). Через крючковидный пучок — связь с амигдалами, темпорополярным произокортексом и передней частью верхней височной извилины. У 10 ПБ нет связей с теменной корой, затылочной корой и нижневисочной корой[5].
Взаимосвязи 10 ПБ были обобщены следующим образом: «в отличие от остальных регионов префронтальной коры оно выглядит не связанным с зонами „нижнего потока“… это единственный префронтальный регион, который преимущественным (и, возможно, исключительным) образом взаимосвязан с надмодальными зонами префронтальной коры, передней височной корой и поясной корой»[2]. Что и обеспечивает высочайший уровень интеграции информации, анализируемой зрительной, слуховой и соматосенсорной системами, позволяя достичь амодальной, абстрактной и концептуальной интерпретации стимулов окружающего мира[5].
Функции
Фронтополярная кора играет важную роль в обеспечении таких высших когнитивных функций, как планирование, принятие решений, осознание и установление логических связей между явлениями/теоретическими положениями, вызов воспоминаний из эпизодической памяти[6].
Этьен Кёклен и Александр Ияфиль полагают, что обеспечение процессов когнитивного разветвления являются главной функцией фронтополярной коры[7]. Когнитивное ветвление позволяет предыдущим выполняемым задачам сохраняться в ожидающем и незаконченном состоянии для последующего их задействования и управления ими после завершения тех когнитивных задач, что обрабатываются в настоящем времени[6]. Наше комплексное поведение и сложная ментальная активность требуют одновременного выполнения множества задач, и предполагается, что передняя префронтальная кора может выполнять генерализующую функцию в организации одновременной работы со многими когнитивными операциями и задачами.
Также следует учитывать взаимовлияние фронтополярной коры и лимбической системы, с которой фронтополярная кора связана через вентромедиальную префронтальную кору.
Примечания
- ↑ Knowlton, Barbara J.; Morrison, Robert G.; Hummel, John E.; Holyoak, Keith J. A neurocomputational system for relational reasoning // Trends in Cognitive Sciences 16 (7). — 2012. — С. 373—381.
- ↑ 1 2 3 Ramnani N, Owen AM. Anterior prefrontal cortex: insights into function from anatomy and neuroimaging // Nat Rev Neurosci. 5(3). — 2004. — С. 184-194. Архивировано 25 июля 2009 года.
- ↑ 1 2 3 4 5 Semendeferi K, Armstrong E, Schleicher A, Zilles K, Van Hoesen GW. Prefrontal cortex in humans and apes: a comparative study of area 10 // Am J Phys Anthropol. 114(3). — 2001. — С. 224-241. Архивировано 26 июня 2017 года.
- ↑ Jacobs B, Schall M, Prather M, Kapler E, Driscoll L, Baca S, Jacobs J, Ford K, Wainwright M, Treml M. Regional dendritic and spine variation in human cerebral cortex: a quantitative golgi study // Cereb Cortex. 11(6). — 2001. — С. 558-571. Архивировано 6 мая 2016 года.
- ↑ 1 2 Petrides M, Pandya DN. Efferent association pathways from the rostral prefrontal cortex in the macaque monkey // J Neurosci. 27(43). — 2007. — С. 11573-11586. Архивировано 26 июня 2016 года.
- ↑ 1 2 Todd S. Braver, Susan R. Bongiolatti. The Role of Frontopolar Cortex in Subgoal Processing during Working Memory // NeuroImage 15. — 2002. — С. 523—536. Архивировано 4 марта 2016 года.
- ↑ Koechlin, E. & Hyafil, A. Anterior prefrontal function and the limits of human-decision making // Science, Vol. 318. — 2007. — С. 594-598.
| Лобная доля |
| ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Теменная доля |
| ||||||||||||||
| Затылочная доля |
| ||||||||||||||
| Височная доля |
| ||||||||||||||
| Междолевые борозды |
| ||||||||||||||
| Сводчатая извилина |
| ||||||||||||||
| Островковая доля |
| ||||||||||||||
| Прочее |
| ||||||||||||||