Цитоскелет

Цитоскеле́т — это клеточный каркас или скелет, находящийся в цитоплазме живой клетки. Он присутствует во всех клетках эукариот, причём в клетках прокариот обнаружены гомологи всех белков цитоскелета эукариот. Цитоскелет — динамичная, изменяющаяся структура, в функции которой входит поддержание и адаптация формы клетки ко внешним воздействиям, экзо- и эндоцитоз, обеспечение движения клетки как целого, активный внутриклеточный транспорт и клеточное деление.

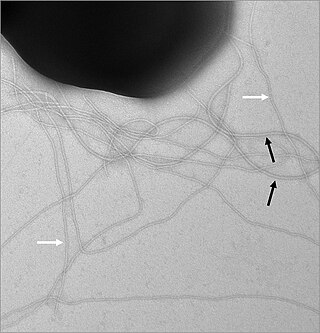
Цитоскелет образован белками, выделяют несколько основных систем, называемых либо по основным структурным элементам, заметным при электронно-микроскопических исследованиях (микрофиламенты, промежуточные филаменты, микротрубочки), либо по основным белкам, входящим в их состав (актин-миозиновая система, кератины, тубулин-динеиновая система).
Цитоскелет эукариот
Клетки эукариот содержат три типа так называемых филаментов. Это супрамолекулярные, протяжённые структуры, состоящие из белков одного типа, сходные с полимерами. Разница заключается в том, что в полимерах связь между мономерами ковалентная, а в филаментах связь составных единиц обеспечивается за счёт слабого нековалентного взаимодействия.
Актиновые филаменты (микрофиламенты)
Порядка 7 нм в диаметре, микрофиламенты представляют собой две цепочки из мономеров актина, закрученные спиралью. В основном они сконцентрированы у внешней мембраны клетки, так как отвечают за форму клетки и способны образовывать выступы на поверхности клетки (ламеллоподии[англ.] и микроворсинки). Также они участвуют в межклеточном взаимодействии (образовании адгезивных контактов), передаче сигналов и, вместе с миозином — в мышечном сокращении. С помощью цитоплазматических миозинов по микрофиламентам может осуществляться везикулярный транспорт.
Промежуточные филаменты (нанофиламенты)
Диаметр промежуточных филаментов составляет от 8 до 12 нанометров. Они состоят из разного рода субъединиц, большинство белков промежуточных филаментов у млекопитающих является кератинами. Для нанофиламентов не характерно присутствие центра организации; они образуют пучки и сетчатые структуры в ядре и в цитоплазме клетки. Промежуточные филаменты обладают высокой эластичностью и обеспечивают устойчивость клетки к механическому и химическому стрессу[1].
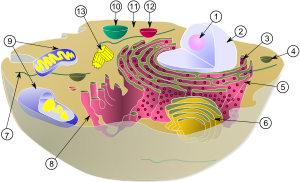
(1) Ядрышко
(2) Ядро
(3) рибосома (маленькие точки)
(4) Везикула
(5) шероховатый эндоплазматический ретикулум (ER)
(6) Аппарат Гольджи
(7) Цитоскелет
(8) Гладкий эндоплазматический ретикулум
(9) Митохондрия
(10) Вакуоль
(11) Цитоплазма
(12) Лизосома
(13) Центриоль и Центросома
Микротрубочки
Микротрубочки представляют собой полые цилиндры порядка 25 нм диаметром, стенки которых составлены из 13 протофиламентов, каждый из которых представляет линейный полимер из димера белка тубулина. Димер состоит из двух субъединиц — альфа- и бета- формы тубулина. Микротрубочки — крайне динамичные структуры, потребляющие ГТФ в процессе полимеризации. Они играют ключевую роль во внутриклеточном транспорте (служат «рельсами», по которым перемещаются молекулярные моторы — кинезин и динеин), образуют основу аксонемы ундулиподий и веретено деления при митозе и мейозе.
Долгое время считалось, что цитоскелетом обладают только эукариоты. Однако с выходом в 2001 году статьи Jones и соавт. (PMID 11290328), описывающей роль бактериальных гомологов актина в клетках Bacillus subtilis, начался период активного изучения элементов бактериального цитоскелета. К настоящему времени найдены бактериальные гомологи всех трех типов элементов цитоскелета эукариот — тубулина, актина и промежуточных филаментов[2]. Также было установлено, что как минимум одна группа белков бактериального цитоскелета, MinD/ParA, не имеет эукариотических аналогов.
Бактериальные гомологи актина
К наиболее изученным актиноподобным компонентам цитоскелета относятся MreB, ParM и MamK.
MreB и его гомологи
Белки MreB и его гомологи являются актиноподобными компонентами цитоскелета бактерий, играющими важную роль в поддержании формы клетки, сегрегации хромосом и организации мембранных структур. Некоторые виды бактерий, такие как Escherichia coli, имеют только один белок MreB, тогда как другие могут иметь два и более MreB-подобных белков. Примером последних служит бактерия Bacillus subtilis, у которой были обнаружены белки MreB, Mbl (MreB-like) и MreBH (MreB homolog).
В геномах E. coli и B. subtilis ген, отвечающий за синтез MreB, находится в одном опероне с генами белков MreC и MreD. Мутации, подавляющие экспрессию данного оперона, приводят к образованию клеток сферической формы с пониженной жизнеспособностью.
Субъединицы белка MreB образуют филаменты, обвивающие палочковидную бактериальную клетку. Они располагаются на внутренней поверхности цитоплазматической мембраны. Филаменты, образуемые MreB, динамичны, постоянно претерпевают полимеризацию и деполимеризацию. Непосредственно перед делением клетки MreB концентрируется в области, в которой будет формироваться перетяжка. Считается, что функцией MreB также является координация синтеза муреина — полимера клеточной стенки.
Гены, отвечающие за синтез гомологов MreB, были обнаружены только у палочковидных бактерий и не были найдены у кокков.
ParM
Белок ParM присутствует в клетках, содержащих малокопийные плазмиды. Его функция заключается в разведении плазмид по полюсам клетки. При этом субъединицы белка формируют филаменты, вытянутые вдоль большой оси палочковидной клетки.
Филамент по своей структуре представляет собой двойную спираль. Рост филаментов, образуемых ParM, возможен с обоих концов, в отличие от актиновых филаментов, растущих только на ±полюсе.
MamK
MamK — это актиноподобный белок Magnetospirillum magneticum, отвечающий за правильное расположение магнитосом. Магнитосомы представляют собой впячивания цитоплазматической мембраны, окружающие частички железа. Филамент MamK выполняет роль направляющей, вдоль которой, одна за другой, располагаются магнитосомы. В отсутствие белка MamK магнитосомы располагаются беспорядочно по поверхности клетки.
Гомологи тубулина
В настоящее время у прокариот найдены два гомолога тубулина: FtsZ и BtubA/B. Как и эукариотический тубулин, эти белки обладают ГТФазной активностью.
FtsZ
Белок FtsZ[англ.] чрезвычайно важен для клеточного деления бактерий, он найден практически у всех эубактерий и архей. Также гомологи этого белка были обнаружены в пластидах эукариот, что является ещё одним подтверждением их симбиотического происхождения.
FtsZ формирует так называемое Z-кольцо, выполняющее роль каркаса для дополнительных белков клеточного деления. Вместе они представляют собой структуру, ответственную за образование перетяжки (септы)[3].
BtubA/B
В отличие от широко распространенного FtsZ, белки BrubA/B[англ.] обнаружены только у бактерий рода Prosthecobacter. Они более близки к тубулину по своему строению, чем FtsZ.
Кресцентин, гомолог белков промежуточных филаментов
Белок был найден в клетках Caulobacter crescentus. Его функцией является придание клеткам C. crescentus формы вибриона. В случае отсутствия экспрессии гена кресцентина клетки C. crescentus приобретают форму палочки. Интересно, что клетки двойных мутантов, кресцентин− и MreB−, имеют сферическую форму.
MinD и ParA
Эти белки не имеют гомологов среди эукариот.
MinD[англ.] отвечает за положение сайта деления у бактерий и пластид. ParA участвует в разделении ДНК по дочерним клеткам.
См. также
- ADF/Кофилин
Примечания
- ↑ Л. Кассимерис. Клетки по Льюину. — пер. 2-го англ. изд. — М.: Лаборатория знаний, 2016. — С. 612-618. — 1056 с. — ISBN 978-5-906828-23-1.
- ↑ Shih Y.-L., Rothfield L. The Bacterial Cytoskeleton. // Microbiology And Molecular Biology Reviews. — 2006. — V. 70., No. 3 — pp. 729—754. PMID 16959967.
- ↑ de Boer PA. Advances in understanding E. coli cell fission (неопр.) // Curr Opin Microbiol.. — 2010. — Т. 13. — С. 730—737. — doi:10.1016/j.mib.2010.09.015. — PMID 20943430.
| Словари и энциклопедии | ||||
|---|---|---|---|---|
Белки цитоскелета | |
|---|---|
| Микрофиламенты |
|
| Промежуточные филаменты | |
| Микротрубочки |
|
| Катенины |
|
| другие |
|