Человек флоресский
| † Человек флоресский | |
|---|---|
 Череп флоресского человека | |
| Научная классификация | |
Домен: Эукариоты Царство: Животные Подцарство: Эуметазои Без ранга: Двусторонне-симметричные Без ранга: Вторичноротые Тип: Хордовые Подтип: Позвоночные Инфратип: Челюстноротые Надкласс: Четвероногие Клада: Амниоты Клада: Синапсиды Класс: Млекопитающие Подкласс: Звери Клада: Эутерии Инфракласс: Плацентарные Магнотряд: Бореоэутерии Надотряд: Грандотряд: Эуархонты Миротряд: Приматообразные Отряд: Приматы Подотряд: Сухоносые приматы Инфраотряд: Обезьянообразные Парвотряд: Узконосые обезьяны Надсемейство: Человекообразные обезьяны Семейство: Гоминиды Подсемейство: Гоминины Триба: Гоминини Подтриба: Хоминина Род: Люди Вид: † Человек флоресский | |
| Международное научное название | |
| Homo floresiensis Brown et al., 2004 | |
| Ареал | |
 | |
| Геохронология | |
Человек фло́ресский (лат. Homo floresiensis) — возможный карликовый вид ископаемых людей. Из-за малого роста флоресский человек известен также как «хоббит» (по аналогии с существами, придуманными Дж. Р. Р. Толкином). Останки Homo floresiensis впервые обнаружены в 2003 году в Индонезии (остров Флорес, пещера Лианг-Буа), где найдены несколько скелетов разной степени сохранности возрастом приблизительно в 13—95 тысяч лет (по последним альтернативным оценкам — 60—100 тысяч лет)[1]. В дальнейшем находки костей карликового размера, возраст которых оценивается в 700 тысяч лет, были сделаны в Мата-Менге поблизости от Лианг-Буа. По единственному известному черепу и наиболее полно сохранившимся скелетам рост флоресских людей оценивается примерно в 1 м, а объём головного мозга в 400 см³, что в три раза меньше объёма мозга современного человека и приближается к параметрам, характерным для австралопитеков. Исследователи отмечают также многочисленные архаичные черты в строении черепа и конечностей флоресских людей.
Вместе с останками флоресских людей были найдены примитивные каменные орудия, обожжённые кости животных и прочие следы материальной культуры. Однако малый рост и в особенности чрезвычайно малый объём мозга вызывает у части научного сообщества сомнения в том, что найденные останки принадлежат отдельному виду людей. Ряд исследователей считает их патологически изменёнными останками человека разумного. Как гипотеза об отдельном виде, так и гипотеза о патологии оставляют открытыми определённые вопросы. В первом случае неясными остаются происхождение карликового флоресского вида, пути его проникновения на остров и время вымирания, во втором — характер патологии, вызвавшей необычные изменения в анатомии.
Открытие и наименование

Останки человека флоресского были впервые обнаружены в 2003 году на индонезийском острове Флорес совместной индонезийско-австралийской археологической экспедицией. Экспедиция под руководством австралийца Майка Морвуда и индонезийца Панджита Суджоно работала в пещере Лианг-Буа на западе острова с 2001 года, продолжая раскопки, которые до этого вёл Суджоно, и постепенно открывая более древние слои отложений. В процессе раскопок в первый год было обнаружено множество костей различных животных, в том числе таких крупных, как стегодоны и комодские драконы; многочисленные грубые палеолитические орудия; и лучевая кость, похожая на человеческую, но маленькая и необычно изогнутая[2]. Находка 2003 года была более значительной, представляя собой неполный скелет возрастом около 18 тысяч лет, предположительно взрослой женской особи очень маленького роста, включавший череп, нижнюю челюсть и большинство зубов. Обнаруженные в сырой пещере кости не были окаменелыми, а, напротив, по фактуре напоминали «картофельное пюре» и «мокрую промокашку», так что в обращении с ними потребовалась предельная аккуратность[3]. Сохранность костей была обеспечена с помощью самодельного препарата, изготовленного местным археологом Томасом Сутикной из моментального клея и жидкости для снятия лака[4].
Лианг-Буа на протяжении тысячелетий служила местному населению местом ритуальных захоронений, для которых была характерна окраска костей охрой и украшение ожерельями из раковин, а позже, с освоением металлов, в захоронениях появились бронзовые и железные орудия труда[5]. Найденный скелет, однако, ничем не напоминал эти позднейшие захоронения: ничто не указывало на ритуальную обработку останков. При этом он хорошо сохранился, вплоть до естественного сочленения костей правой ноги и таза. Участники экспедиции сделали вывод, что вскоре после смерти тело погрузилось в жидкую грязь мелкого водоёма, что защитило останки от растаскивания зверями; подобным образом сохранились известные останки Люси (женской особи австралопитека афарского возрастом свыше 3 млн лет) и мальчика из Турканы (человека работающего возрастом около 1,6 млн лет)[6].
В 2004 году обнаружены дополнительные останки, по которым исследователи восстановили полный внешний облик неизвестного гоминина и время существования предполагаемой популяции — 12—95 тысяч лет назад (позже верхняя граница этого периода, которую задаёт слой вулканического туфа, уточнялась до 13 100[7], а затем до 17 000 лет[8], а в 2016 году были опубликованы новые оценки, основанные на радиоизотопном анализе останков и датирующие их периодом между 60 и 100 тысячами лет назад[1]). Также вместе с останками были обнаружены каменные орудия, указывающие на способности к охоте и строительству[9][10].
После того, как был сделан вывод о том, что открыт новый биологический вид, гомининам метрового роста, первоначально получившим от исследователей прозвище «хоббиты», было предложено присвоить видовое имя Homo hobbitus. Высказывалось также мнение, что речь идёт о новом роде, для которого предлагалось имя Sundantropus — эту точку зрения отстаивал приглашённый из Австралии палеоантрополог Питер Браун, убеждённый, что физические характеристики останков (в частности, объём черепной коробки) не позволяют отнести их к роду Homo. Лишь после долгих споров другим исследователям и рецензентам удалось убедить Брауна в том, что череп первой найденной особи принадлежит представителю рода Homo, и вид получил имя «человек флоресский» (лат. Homo floresiensis)[11]. Впервые сообщение о нём в научной прессе появилось в октябре 2004 года[12] в журнале Nature после того, как исследовательская группа успешно хранила сведения об открытии в секрете больше года; такие шаги были связаны с тем, что преждевременное появление новостей о находке в прессе вызвала бы отмену публикации материалов в Nature — одном из авторитетнейших научных журналов[13]. Сразу после выхода номера журнала со статьями о находке на Флоресе эта новость была растиражирована другими изданиями: по оценке Майка Морвуда, материалы о флоресском человеке появились в семи тысячах газет и новостных журналов и почти на ста тысячах сайтов. Вскоре журнал National Geographic, пользующийся высокой репутацией, но значительно более общедоступный, чем Nature, начал работу над целой серией материалов о «хоббите», включая съёмку документального фильма для одноимённого телеканала[14].
В конце 2004 года индонезийский профессор-палеоантрополог Теуку Якоб, ведущий специалист в этой области у себя в стране, забрал большую часть костных останков, связываемых с человеком флоресским, из Национального археологического исследовательского центра в Джакарте. Это было сделано с разрешения одного из руководителей проекта — индонезийца Панджи Суджоно[англ.], которого с Якобом связывала дружба со времён освободительной борьбы против японских оккупантов[15], и вопреки возражениям австралийского археолога Майка Морвуда, осуществлявшего общую координацию экспедиции[16][17].
После этого останки находились в полном распоряжении Якоба на протяжении трёх месяцев, что вызвало возмущение остальных учёных, с точки зрения которых это означало монополию на останки одной исследовательской группы. После разразившегося скандала Якоб, один из ведущих противников теории об особом месте человека флоресского в биологической классификации, вынужден был вернуть кости, но многие из них были возвращены с повреждениями: на одних были свежие надрезы, другие были поломаны и кое-как склеены снова. Работник лаборатории Якоба сообщил, что хрупкие кости были повреждены при попытках сделать их слепки[4]. Другие кости, в частности тазовые, по-видимому, пострадали при транспортировке[16] (по словам Якоба, тогда же пострадали и остальные кости[18]). Две кости ноги LB1 остались у Якоба для дальнейших исследований. Возмущение самовольными действиями Якоба было настолько сильным, что гневные письма получал даже сотрудник лейпцигского Института Макса Планка Жан-Жак Юблен, для исследований возможной ДНК использовавший однограммовый скол с одного из рёбер LB1. По мнению Юблена, Якоб подвергся несправедливым нападкам, так как западная пресса выставила его в виде вора, похитившего чужое открытие[19]. Юблен заявил, что поведение австралийских археологов представляет собой «чистейший образец научного неоколониализма и западной заносчивости»[20].
После скандала индонезийские власти запретили археологам в 2005 году доступ к пещере Лианг-Буа. Это вызвало обвинения в попытке спасти научный авторитет профессора Якоба, считающегося «королём индонезийской палеоантропологии», так как новые находки могли доказать его неправоту. В 2007 году было получено разрешение на продолжение раскопок[18]. В конце 2015 года было сообщено, что международная экспедиция ведёт раскопки в обнаруженной ещё в 2006 году пещере Лианг-Бавах, которая связана проходом с Лианг-Буа и в которой сохранились более древние отложения; авторы статьи предполагают, что эта вторая пещера служила обитателям Лианг-Буа складом и путём к бегству. К моменту публикации были исследованы лишь наиболее поздние слои, содержащие только свидетельства бытовой деятельности людей современного типа, однако авторы выражали надежду, что более ранние отложения позволят узнать больше о человеке флоресском[21].
В 2024 году было сообщено, что на раскопках Мата-Менге, в том же регионе, что и Лианг-Буа, были обнаружены челюсть, восемь зубов и обломки плечевой кости, принадлежащие взрослым гомининам, имевшим ещё меньшие размеры, чем найденные в Лианг-Буа. Возраст находок был оценён в 700 тысяч лет[22]
Анатомия
Отличительными чертами строения тела человека флоресского являются общий маленький рост и маленький размер черепной коробки. От человека разумного его также отличают общая форма черепа (более близкая к Homo ergaster, чем к человеку разумному), отсутствие подбородочного выступа, отличающего человека разумного от более древних гоминидов, приплюснутый нос и мощные надбровные дуги, также характерные для ископаемых видов рода Homo[23]. По строению черепа и конечностей человек флоресский также принципиально отличается от всех известных популяций Homo erectus, а по соотношению длины отдельных костей — и от Homo ergaster, сближаясь с ещё более примитивными формами — такими, как австралопитеки.
Размеры тела
Голотипом для предполагаемого нового вида стала первая, лучше всего сохранившаяся особь, по номенклатуре LB1 и по прозвищу Фло[4]. Это практически полный скелет половозрелой (ориентировочно 30-летнего возраста) женской особи ростом, по разным оценкам, от 106[9] до 108 сантиметров (оценка роста основана на размерах бедренной кости) или немного ниже, с учётом низкого черепа[24]. Для второй относительно хорошо сохранившейся особи, LB8, рост оценивался в 109 сантиметров на основании измерения большеберцовой кости, поскольку у этого экземпляра бедренные кости отсутствуют[9]; однако большеберцовая кость LB8 на 2 см короче, чем аналогичная кость Фло, и в дальнейшем была сделана новая оценка, основанная на известных пропорциях тела LB1, согласно которой рост LB8 был даже меньше, чем у Фло[24]. Этот рост находится далеко за пределами нормального распределения роста для современного человека разумного[25] и значительно меньше среднего роста женщин-андаманок (137 см), представляющих, наряду с пигмеями мбути, самые низкорослые современные племена[26].
В целом, человек флоресский был предположительно меньше не только здоровых представителей любой современной человеческой расы, но и Homo erectus и известных видов австралопитеков[27]. В литературе подчёркивается, что оценки делались на основании костного материала женской особи; неизвестно, есть ли среди найденных в пещере Лианг-Буа останков мужские, и неизвестно также, был ли характерен для флоресских людей половой диморфизм, при котором мужчины были бы существенно крупней женщин[24].
Масса тела, которую первые исследователи оценивали исходя из его параметров, которые они посчитали более близкими к шимпанзе, чем к человеку разумному[9], должна была составить предположительно 16—29 килограммов[28]. За последующие годы исследований флоресских находок было предложено несколько оценок как роста наиболее хорошо сохранившихся экземпляров (мало отличающихся одна от другой), так и массы тела (демонстрирующих более значительные расхождения). Оценки массы тела базировались на различных параметрах бедренной и большеберцовой костей, а также соединяющего кости ноги сустава и составляли от 25 до 35 кг, с медианным значением около 32,5 кг. Если предполагаемый рост Фло и LB8 меньше, чем у любых известных низкорослых народов, то оцениваемая масса тела находится в пределах допустимого для современных пигмеев диапазона, а индекс массы тела и весовой индекс (рассчитываемый как соотношение кубического корня массы в килограммах к росту в метрах), напротив, существенно выше. Таким образом, телосложение флоресских людей было более массивным, чем у наших современников, напоминая скорее телосложение ранних гоминин — таких, как австралопитек афарский — и небольших африканских человекообразных обезьян (бонобо и швейнфуртовский шимпанзе)[24].
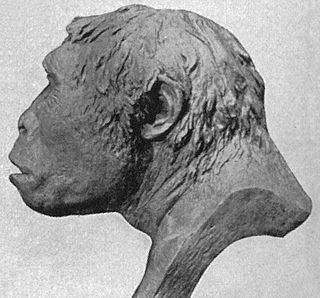
Череп и мозг

Единственный найденный до настоящего времени в пещере Лианг-Буа череп принадлежит Фло. Уже при находке черепа исследователи обратили внимание на такие черты, как небольшая (возможно, посмертная) асимметрия правой челюстной дуги и правого теменного края лобной кости; правый венечный шов на черепе разошёлся. Более поздние повреждения (уже в процессе раскопок) получила левая сторона черепа, в частности передние лицевые кости и теменная кость в районе брегмы (точки, соответствующей месту соединения сагиттального и венечного швов). Тем не менее череп в целом достаточно хорошо сохранился, позволив сделать ряд выводов о внешности Фло. В частности, полностью закрывшиеся швы на своде и основании черепа показывают, что Фло к моменту смерти была взрослой, полностью сформировавшейся особью[24] (об этом же свидетельствуют износ зубов и прорезавшиеся третьи моляры[29]).
Череп LB1, по-видимому, вмещал чрезвычайно маленький по объёму головной мозг. Объём полости черепа составлял 417 см³[30], что значительно уступает не только современному человеку, но и черепной коробке человека прямоходящего, для которого размер мозга составлял от 800 до 1250 см³. Такой объём сопоставим с объёмом известных слепков полости черепа австралопитеков, который варьировал от 427 до 545 см³[31]. Другие морфологические характеристики черепа человека флоресского при рассмотрении в совокупности резко отличают его как от типичного черепа современного человека, так и от черепа австралопитеков. В то же время он обладает рядом сходных черт с черепами человека прямоходящего (в большей степени для экземпляров не азиатского происхождения) и человека умелого[32]. Малые размеры черепа Фло напоминают о древнейших австралопитеках, но в остальном он не похож на их массивные, имеющие тяжёлые мощные челюсти черепа, а его лицевой отдел, по мнению одного из исследователей останков из пещеры Лианг-Буа, Уильяма Юнгерса, скорей напоминает лица ранних представителей рода Homo[24].
Низкий, широкий и длинный свод черепа LB1, по очертаниям напоминающий черепа́ человека прямоходящего, достигает максимальной ширины достаточно низко — сразу над сосцевидными отростками височной кости. Угол основания черепа достаточно крутой, порядка 130°, что больше, чем не только у современных людей, но и у большинства ископаемых гоминин. На затылке расположен слаборазвитый поперечный гребень, но затылочный бугор развит меньше, чем у человека прямоходящего, а теменной гребень, характерный для последнего, на черепе Фло, по-видимому, отсутствует. В левой затылочно-теменной области отмечено асимметричное уплощение, которое исследователи рассматривают как результат затылочной позиционной плагиоцефалии. Кости свода черепа толстые, что встречается как у человека прямоходящего, так и у современных людей, и содержат многочисленные воздушные полости, в частности, в височных костях, в губчатом веществе костей свода и костях надглазничной области[24]. Микротомографическое исследование, опубликованное в начале 2016 года, показывает, что расположение воздушных полостей (в том числе их отсутствие во фронтальной части черепа) и толщина костей черепа укладываются в известные рамки для примитивных гоминидов (в частности человека прямоходящего), но не демонстрируют прогрессивных черт, свойственных для человека разумного[33].
Выпуклые надбровные дуги черепа LB1 не образуют тем не менее сплошного валика; не отмечена также характерная для человека прямоходящего треугольная форма дуг, выступающих за контуры лица. Лобная кость плавно уходит назад, увенчана невысоким сагиттальным гребнем и отделена от надбровных дуг вдавленной зоной, известной как надглазничный жёлоб. Шиловидные отростки височных костей, по-видимому, отсутствовали изначально, а не были утеряны после смерти Фло. Относительная высота лица меньше, чем у австралопитеков, а жевательные мышцы не настолько сильно развиты[24].
Некоторые характеристики мозга Фло приближают его к мозгу человека прямоходящего, в то время как другие (удлинённая затылочная доля коры полушарий, выступающая над мозжечком, возможно, связанная с вытеснением отделов, ответственных за зрение, более развитыми ассоциативными зонами[34], а также отсутствие лобно-глазничной борозды (лат. sulcus fronto-orbitalis), исчезающей в процессе эволюции приматов по мере того, как развиваются лобные доли коры[35]) — к головному мозгу человека разумного. Головной мозг Фло также отличает ряд особенностей, не имеющих аналогов среди других приматов. Одной из таких особенностей являются чрезвычайно развитые (относительно общего объёма мозга) височные доли. Этот отдел мозга у современного человека отвечает в частности за опознание предметов и лиц[36]. Другой отличительной чертой были две мощные извилины в 10-м поле Бродмана, которые, по мнению создававшей виртуальный эндокран учёной-нейроантрополога Дин Фальк[англ.], потенциально указывают на хорошо развитое воображение и способности к абстрактному мышлению и планированию[37].
Нижняя челюсть и зубы
В отличие от других частей черепа, при раскопках в Лианг-Буа были обнаружены две хорошо сохранившиеся нижние челюсти. В обоих случаях они демонстрируют ряд примитивных характеристик, некоторые из которых (заметные режущие кромки на клыках и наличие нёбных дужек по обе стороны носового отверстия) роднят их с челюстями автралопитеков. Челюсти из Лианг-Буа отличаются очень крупными относительно общих размеров тела зубами, сильно выраженной кривой Шпее (строением, при котором жевательные поверхности зубов нижней челюсти образуют вогнутую сагиттальную кривую) и выраженными бугристостями для крепления жевательных мышц. Нехарактерными для современного человека являются полное отсутствие подбородочного выступа (хотя, как указано в разделе Гипотезы о «больных хоббитах», такое явление встречается у представителей австраломеланезийской расы[38]), сильно утолщённый подбородочный симфиз, к которому с внутренней и наружной стороны прилегают дополнительные валики, также знакомые по челюстям австралопитеков[24]. Имеются также черты, отмеченные у ранних представителей рода Homo, в том числе питекантропа и дманисийского человека[39].
Зубы из Лианг-Буа сильно изношены, местами до плоской жевательной поверхности и обнажения дентина, что характерно для культур охотников и собирателей; износ нижних зубов LB1 асимметричен, становясь более сильным к левым коренным зубам, что, вероятно, связано с неправильным прикусом из-за затылочной позиционной плагиоцефалии. На зубах этой челюсти обнаружен зубной камень, но нет никаких признаков кариеса, как и на второй обнаруженной челюсти[24]. Анализ зубов не менее чем четырёх отдельных особей (LB1, LB6, LB15 и LB2), результаты которого были опубликованы в конце 2015 года, показывает, что для клыков и премоляров из Лианг-Буа характерно примитивное строение, напоминающее строение аналогичных зубов человека прямоходящего. При этом моляры демонстрируют высокоразвитую морфологию, в некоторых отношениях превосходящую даже зубы человека разумного. Примером последней может служить сильно уменьшенный первый коренной зуб — черта, обычно увязываемая с высокими технологиями обработки пищи, но в условиях примитивной культуры Лианг-Буа остающаяся необъяснимой. Авторы публикации делают вывод, что флоресские останки принадлежат отдельному виду, по прямой линии восходящему к азиатским Homo erectus (в частности, явантропам), а не эволюционировавшему параллельно им[40].
Более древние зубы из раскопа Мата-Менге демонстрируют характеристики, роднящие их с зубами человека прямоходящего. Верхний клык отличают примитивные черты, схожие не только с зубами представителей человека прямоходящего из Сангирана (Индонезия), но и с зубами более древних австралопитеков; специфический износ этого зуба указывает на примитивное строение прилегающего к нему первого моляра. Третий моляр из той же челюсти достаточно близок по устройству к молярам человека прямоходящего из Сангирана, а также имеет определённое сходство с зубами Homo ergaster, но более прогрессивен, чем моляры человека умелого, и в целом ближе к зубам человека разумного, чем к ним[41].
Конечности

За время исследования флоресских останков отмечались такие нехарактерные для современного человека особенности, как малый изгиб костей рук и толщина костей ног. Хотя эти характеристики подтверждаются оппонентами, их значение для выделения флоресского человека в отдельный от Homo sapiens биологический вид оспаривается[42].
Плечевая кость очень мощная, но без выраженных мест крепления мышц; в отличие от плечевой, не менее мощная локтевая кость демонстрирует следы крепления мышц в районе соединения с плечевой[24]. У плечевой кости, найденной в Мата-Менге, отсутствует характерная искривлённость ствола, указывающая на способности к древолазанию и характерная для самых ранних гоминин, что мможет свидетельствовать о достаточно поздней эволюции вида[22]. Исследование плеча и предплечья флоресского человека в 2007 году показало, что предплечье повёрнуто на 120° по отношению к локтевому суставу. У современного человека угол поворота составляет 145—165°. Недостаточный изгиб, который мог бы помешать процессу изготовления орудий, однако, искупается более короткой и вывернутой вперёд ключицей, создающей впечатление пожатия плечами. За счёт этого Фло могла изгибать локоть так же, как современные люди[43]. Хорошо сохранившаяся лопатка LB6 по строению и положению соответствует устройству плеча Фло — она удлинена и развёрнута так, что оказывается сбоку от грудной клетки, суставной впадиной вперёд, что может указывать на форму груди, отличную от бочкообразной груди современных людей. В то же время ряд характеристик лопатки, в том числе суставный угол, указывающий на ориентацию плечевого сустава, и близкая к горизонтальной ориентация ости, напоминает строение лопаток современного человека[24].
Также в 2007 году исследование трёх костей запястья, найденных на Флоресе в 2003 году и приписываемых LB1, показало их архаичный характер. У этих костей отсутствуют характерные для современного человека разумного и неандертальца прогрессивные черты, и по строению они напоминают аналогичные кости современных человекообразных обезьян, а также древнейших ископаемых гоминидов — австралопитеков и человека умелого. Исследователи пришли к выводу, что эти примитивные черты не могут быть результатом какой-либо из известных болезней или аномалией развития[44]. Один из ведущих оппонентов гипотезы об отдельном виде, доктор Алан Торн, со своей стороны указал, что принадлежность всех костей, найденных разрозненными, одной особи не доказана; по его мнению, отличия этих костей от костей современного человека не больше тех, которые возможны в пределах одного вида[45].
Строение и пропорции таза человека флоресского мало отличаются от строения таза других высокоразвитых гоминин, в том числе и человека разумного. Проксимальный отдел бедра Фло, однако, в дополнение к прогрессивным демонстрирует некоторые архаичные черты, напоминающие об австралопитеках — относительно маленькую головку и длинную, сплюснутую в антеропостериорном плане шейку. Бедренная кость, массивная, как и кости руки, очень короткая относительно других размеров тела (всего 280 мм). Также толстой и короткой является и большеберцовая кость, овальная в сечении, что отмечено у австралопитеков, но встречается, хотя довольно редко, и у современных людей; пропорции малоберцовых костей вполне современные. Наиболее архаичной частью нижних конечностей являются ступни, для которых характерны короткие большие пальцы и отсутствие выраженного продольного свода стопы — черта, описанная у австралопитеков. Согласно публикациям конца 2000 — начала 2010-х годов, длина стопы также очень велика относительно длины бедра и ноги в целом (схожие пропорции встречаются у бонобо), что в основном связано с малой длиной бедренной кости[24]. Однако выводы о большой относительно роста длине ступней были оспорены десятью годами позже; авторы нового исследования высказали предположение, что кости нескольких разных особей были приписаны одной, что привело к искажённым представлениям о пропорциях тела флоресских людей[46]. В целом нижние конечности явно приспособлены для передвижения на двух ногах, но лишены некоторых прогрессивных черт, характерных для современного человека[24]; из наличествующих прогрессивных черт можно отметить прилегающий (а не противостоящий) большой палец[47].
Соотношение длин плечевой и бедренной костей, известное как плечебедренный индекс, у LB1 намного выше, чем не только у человека разумного, но и у человека прямоходящего, и ближе всего к таковому у Люси — наиболее хорошо сохранившейся особи австралопитека афарского[48]. Соотношение длины лучевой кости к длине бедренной кости (67,9 %) также нехарактерно для современного человека, для которого наибольшее значение этого соотношения составляет чуть больше 60 %; аналогичное соотношение меньше даже у такого архаичного вида, как человек работающий, и наиболее близкие величины наблюдаются у австралопитека гари[49]. При этом соотношение длин лучевой и плечевой костей и соотношение длин большеберцовой и бедренной костей, составляющие у Фло соответственно 78 % и 84 %, вполне современны и отличаются от зафиксированных у австралопитеков[24].
Попытки реконструкции облика

После открытия в Лианг-Буа регулярно совершаются попытки реконструкции облика флоресских людей. Уже первая публикация сопровождалась графической реконструкцией, созданной художником Петером Схутеном; интересно, что рисунок Схутена изображал мужчину-охотника, в то время как единственный известный череп и наиболее полный скелет (LB1) принадлежат женской особи[51]. Среди авторов, реконструировавших облик Фло, также французский скульптор Элизабет Дейне[фр.], создавшая целый ряд скульптурных портретов ископаемых гоминидов. Скульптурная ростовая реконструкция «маленькой женщины с Флореса», созданная Дейне, экспонируется с 2007 года в Музее человека в Париже[52]. Художественные реконструкции создавались также студией палеохудожницы Карен Карр и Вуллонгонгским университетом (сотрудником которого является Майк Морвуд — один из первооткрывателей «хоббита» с Флореса[53]). Ещё один портрет флоресского человека был размещён в журнале Science в 2009 году[54].
В 2012 году на ежегодной Австралийской археологической конференции доктор Сьюзен Хейз представила реконструкцию лица Фло, выполненную с использованием методов судебной медицины[55]. Корреспондент Scientific American отмечала, что полученная реконструкция близка к облику человека разумного[56]. В 2013 году команда исследователей из Университета штата Нью-Йорк в Стоуни-Брук, Центра эволюции человека и древней окружающей среды в Шенкенберге (Германия) и Университета Миннесоты провела подробный 3D-морфологический анализ черепа Homo floresiensis и пришла к выводу о его близости к человеку прямоходящему. Гипотезу патологии «следует признать несостоятельной», — говорится в выводах исследования[57].
Помимо облика, делаются попытки реконструкции моторики Homo floresiensis, однако в условиях ограниченного материала выводы исследователей могут серьёзно отличаться друг от друга. Так, в 2008 году увидела свет публикация на основе анализа таза и нижних конечностей Фло, в которой был сделан вывод, что походка флоресских людей была похожа на походку современного человека[58]. Однако практически одновременно на встрече Американской ассоциации физических антропологов был представлен доклад, в котором говорилось, что Фло с её плоскими и длинными относительно роста ступнями высоко задирала ноги при ходьбе и в целом передвигалась медленно и неуклюже, подобно клоуну в его огромных башмаках; бегать с такими ногами нельзя ни на короткую, ни на длинную дистанцию[59].
Материальная культура
Ещё до раскопок в пещере Лианг-Буа Флорес был известен как место обнаружения древних каменных орудий. Возраст простейших каменных инструментов и отщепов, найденных при раскопках в пещере Мата-Менге, расположенной к востоку от Лианг-Буа, оценивается в 880—900 тысяч лет. Позже подобные примитивные орудия были найдены неподалёку от Мата-Менге, на выпасе Воло-Сеге, и датированы приблизительно миллионом лет назад. Этим находкам сопутствовали останки некоторых видов местной фауны, в том числе карликового стегодона S. florensis florensis[60]. Таким образом, установлено, что на Флорес гоминины прибыли впервые не позднее, чем миллион лет назад, хотя связь этих древних гоминин и человека флоресского не доказана[61]. В то же время через несколько лет после первых сообщений о находках в пещере Лианг-Буа, содержавших заявления о «прогрессивных» каменных орудиях, появились исследования, показывающие, что эти орудия на самом деле достаточно грубы. По технике изготовления они ближе к находкам из Мата-Менге и другим орудиям эпохи палеолита из Юго-Восточной Азии, чем к известным по другим регионам орудиям человека разумного[62].
В пещере Лианг-Буа к 2009 году было найдено свыше 11 тысяч каменных орудий, возраст самых древних из которых составляет порядка 95 тысяч лет, а самых новых — около 12 тысяч лет. Как и кости гоминин, каменные орудия в пещере не найдены в слоях более поздних, чем вулканический туф, датируемый двенадцатью тысячами лет назад. В целом можно выделить среди найденных в пещере камней булыжники-«заготовки», «молотки» — небольшие продолговатые камни вулканического происхождения с большими переплетёнными кристаллами и следами долгого использования на рабочей поверхности — и изготовленные с их помощью орудия. Среди последних преобладают простые отщепы, отколотые от больших кусков камня, значительно меньше процент орудий, прошедших вторичную обработку для уменьшения размера или придания остроты. Техника изготовления орудий Лианг-Буа практически не менялась на протяжении всего периода. В более поздних голоценовых слоях в этом же районе найдены каменные орудия, связанные с захоронениями людей современного типа и изготовленные из более качественного камня — на 60 % эти находки выполнены из кремнистого сланца, тогда как ниже слоя вулканического туфа таких орудий только 17 %. Кроме того, среди поздних находок чаще встречаются орудия со следами огня, а также отполированные образцы. Однако в целом технология изготовления и этих поздних орудий не отличается от той, что использовалась 95 тысяч лет назад[63].

По мнению Марка Мура, одного из первооткрывателей останков в Лианг-Буа, сходство техники изготовления орудий ниже и выше вулканического слоя может означать, что первые Homo sapiens, прибывшие на Флорес, встречались с его более древними обитателями и переняли у них эту технику. В то же время эти орудия настолько просты в изготовлении, что в своё время в экспериментальном порядке удалось обучить ему даже бонобо. По уровню сложности они не превосходят наиболее примитивные образцы олдувайской культуры, возникшей более двух миллионов лет назад, и совпадение форм может быть результатом конвергенции, связанной со сходными нуждами более раннего и более позднего населения острова. Это, в свою очередь, позволило археологу Джону Ши предположить, что и более ранние образцы — возрастом до 45 тысяч лет — могли быть делом рук человека разумного, который появился в Индонезии в это время[64]. Следует, однако, учитывать, что археологические находки в более поздних слоях указывают на принципиально более высокий уровень культурного развития. Помимо практически не встречающихся в более ранний период следов использования огня, в этих слоях обнаружены примитивные украшения, красители и свидетельства культовых захоронений[65]. В более поздних отложениях встречаются раковины, попавшие в этот район издалека, а также свидетельства раннего скотоводства — человек разумный завёз на Флорес свиней и крупный рогатый скот[66].
Обитатели пещеры Лианг-Буа были, по всей видимости, всеядными охотниками и собирателями. На состав их рациона указывают найденные в пещере многочисленные костные останки, включающие кости рептилий (в том числе молодых стегодонов, комодосских драконов, птиц и местных гигантских крыс), а также растительная органика. Среди кухонных отбросов встречаются обожжённые кости и древесный уголь (хотя и в меньшем количестве, чем в более поздних отложениях), а расположенные по кругу камни со следами огня могут быть остатками кострища (в 2016 году было объявлено, что в Лианг-Буа найден и очаг, по всей видимости сложенный уже человеком разумным около 41 тысячи лет назад, то есть намного ближе к предполагаемому времени исчезновения флоресских людей, чем предыдущие находки[67]). То, что в пещере найдены кости крупных животных, послужило основанием для предположений о том, что флоресский человек охотился группами, что в свою очередь может означать использование речевой коммуникации[68].
Проблема классификации
С момента сообщения о находке в Лианг-Буа сформировались два основных взгляда на то, кому принадлежат найденные останки. Сторонники первого взгляда рассматривают их как принадлежащие представителям карликового вида Homo, сформировавшегося в условиях островной изоляции (среди учёных, разделяющих эту точку зрения существуют разногласия относительно предков этого вида и времени его окончательного исчезновения). Вторая школа трактует останки как принадлежащие изуродованным патологией современным людям (Homo sapiens); в разное время и разными исследователями выдвигались различные гипотезы о характере патологий, сформировавших облик Фло и других найденных особей. Все эти теории Дин Фальк — одна из главных апологетов теории об отдельном виде Homo floresiensis — называет «гипотезами о больных хоббитах»[69]. Предпринятые в 2006 году попытки вычленить ДНК из одного из зубов, найденных в Лианг-Буа тремя годами раньше, не принесли успеха, как предполагается, из-за того, что при температурах, развивающихся при работе бормашины на скорости 1000 оборотов в минуту, ДНК быстро разрушается, а также из-за того, что материалом для извлечения ДНК служил дентин, а не цемент[70]. Окончательные выводы в отсутствие дополнительных черепов или образца ДНК (либо до обнаружения современной патологии, диагноз которой совпадает с известными чертами останков из Лианг-Буа) пока не сделаны[31][71][72].
Гипотеза о карликовом виде людей

Начиная с первой публикации в научной прессе в 2004 году существует популярная точка зрения, согласно которой находка в пещере Лианг-Буа представляет собой останки ранее неизвестного вида людей — Homo floresiensis, эволюционировавшего параллельно современному человеку разумному[12]. По росту Фло — единственный экземпляр с Флореса, для которого обнаружен относительно полный скелет, — схожа с Люси, известной представительницей вида Australopithecus afarensis, но объём её черепа меньше, чем у любого известного черепа австралопитеков[73]. В качестве объяснения малому росту и размеру головы представителей предполагаемого вида предлагается гипотеза, согласно которой развитые приматы, как и другие биологические виды, подвержены эффекту островной карликовости — уменьшения физических размеров в условиях долгой изоляции и отсутствия крупных хищников[28][74]. Известны случаи измельчания в островных условиях за относительно короткие сроки даже человека разумного: так, на том же Флоресе средний рост останков Homo sapiens возрастом 7000 лет существенно больше, чем останков возрастом от 3 до 3,5 тысяч лет[75].
Сторонники теории отдельного вида расходятся в вопросах о его происхождении и времени окончательного исчезновения. Предполагаемый вид мог развиться как на самом Флоресе (как указано выше, находки обработанных орудий труда позволяют датировать появление древних гоминин на острове не позднее чем 1 млн лет назад), но не исключена и возможность, что его предки могли уже прийти на Флорес в таком виде[12]. Хотя ранние публикации рассматривали человека флоресского как потомка питекантропа, новые исследования указывают на различия, не позволяющие считать азиатского человека прямоходящего предком карликов с Флореса: примитивные черты строения черепа и рук указывают на родство с более ранними гомининами, такими, как австралопитеки и человек умелый[32][44][76]. Тазовый пояс единственного относительно полного скелета из Лианг-Буа, LB1 (Фло), шире относительно роста, а бедренные кости отклоняются меньше, чем у представителей рода Homo, что может означать меньшую приспособленность к прямохождению[73]. Левая кисть Фло по строению ближе всего к останкам африканских гоминин возрастом не моложе 1,7 млн лет[76]; в то же время конструкция плечевой кости из Мата-Менге более прогрессивна и указывает на неприспособленность к жизни на деревьях[22]. Авторы опубликованной в 2024 году статьи пишут о большем сходстве плечевой кости с костями Homo naledi, чем более раннего человека умелого[41]. Строение черепа и в особенности челюстей и зубов тоже значительно более современно, чем у австралопитеков, и ближе к человеческому. Большое затылочное отверстие у́же, чем у австралопитеков, сосцевидные отростки височной кости более глубокие и толстые, основание черепа расположено под большим углом, а высота черепной коробки больше, чем у австралопитеков. Относительно общих размеров тела зубы LB1 крупней, чем у человека разумного или человека работающего, но не крупней, чем у человека умелого. Флоресские останки демонстрируют ряд черт, объединяющих их с известными останками человека прямоходящего, а некоторые черты даже более современны — в частности, у черепа Фло отсутствуют характерные для Homo erectus мощные надбровные дуги[73].
Австралийский специалист по доисторической эпохе Роберт Беднарик отмечает, что сам факт неспособности антропологов прийти к единым выводам о том, кем является флоресский человек — измельчавшим человеком прямоходящим, потомком человека умелого или австралопитеком, — заставляет задуматься о несостоятельности современной палеоантропологии[77].
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Отдельные характеристики найденных на Флоресе костей, однако, не схожи ни с австралопитеками, ни с человеком прямоходящим и более поздними представителями рода Homo. В частности, плечевой комплекс Фло обнаруживает сходство с примитивным устройством плеча у останков человека работающего, возраст которых составляет 1,5 млн лет. При попытке провести кладистический анализ в 2009 году на основе 60 признаков (в основном связанных со строением черепа и нижней челюсти) были получены два возможных кладистических дерева: согласно одному из них флоресский человек ответвился от основного эволюционного ствола позже человека рудольфского в позднем плиоцене, а согласно второму — позже человека умелого в раннем плейстоцене. Оба дерева позволяют вычленить большую статистически значимую кладу, в которую человек флоресский входит вместе с человеком умелым, человеком прямоходящим, человеком работающим, родезийским человеком и гоминидом из Дманиси[76]. Последнего вскоре после этой публикации российский палеонтолог Александр Марков назвал возможным предком флоресского человека[81], хотя в кладистическом анализе дерево, предполагающее такую эволюцию, названо менее вероятным[76], а австралийский палеоантрополог Питер Браун считает более прогрессивные черты, характерные для человека из Дманиси, доказательством того, что предки флоресских людей ушли из Африки раньше, чем предки гоминин из Грузии, и, возможно, даже раньше, чем оформился род Homo[82]. В 2015 году были опубликованы результаты байесовского анализа, в котором использовались более 300 морфологических характеристик ископаемых гоминин; анализ показал близость характеристик H. floresiensis с аналогичными чертами австралопитека седиба и дманисийского гоминида, и авторы публикации также пришли к выводу, что предки флоресского человека, схожие с человеком умелым, покинули Африку до появления человека прямоходящего, возможно, даже став первыми гомининами, сделавшими это, и эволюционировали дальше уже в Азии[80]. Через два года исследовательская группа из Австралийского национального университета, проводившая сравнительный анализ по 133 параметрам доступных костных останков, также пришла к выводу, что гипотеза о происхождении флоресских гоминидов от человека прямоходящего несостоятельна и данные указывают на близкое родство с человеком умелым[83].
О новой находке на Флоресе, возможно, способной пролить свет на эволюцию флоресских людей, было сообщено в середине 2016 года: при раскопках в пещере Мата-Менге, где ранее были найдены каменные орудия возрастом около миллиона лет, в 2014 году были обнаружены обломок челюсти и принадлежащие трём разным особям шесть зубов, по размерам и морфологии напоминающие известные останки из пещеры Лианг-Буа (один из зубов демонстрирует ещё более примитивные черты). Возраст останков из Мата-Менге с помощью различных методов датировки оценивается примерно в 700 тысяч лет[84]. Эти кости, как и найденные чуть позже в том же районе обломки плечевой кости, по оценке исследователей, указывают на происхождение человека флоресского от ранних индонезийских популяций человека прямоходящего, а не от более ранних видов Homo или австралопитеков[41].
Нерешённым остаётся также вопрос о том, как примитивный вид людей — или даже ещё более архаичных гоминин — попал на Флорес, который отделяли от других островов Сундского шельфа не менее 19 км водной поверхности даже в период, когда уровень моря был на 100 метров ниже, чем в наше время (остров расположен восточней линии Уоллеса-Хаксли, очерчивающей пределы континентального шельфа Индонезии[85], хотя выдвигалась гипотеза о перешейке, существовавшем между Флоресом и соседним крупным островом Сулавеси). Если предком флоресского человека был человек прямоходящий, то, возможно, следует рассмотреть гипотезу, что около миллиона лет назад представители этого вида были уже достаточно развиты, чтобы строить плоты или другие средства передвижения по воде. Существуют исследования, показывающие возможность постройки лодки с использованием палеолитических каменных орудий, причём одна из таких лодок с экипажем из пяти человек прошла тысячу километров морем от Тимора до Австралии[61]. В рамках съёмок фильма для National Geographic несколько исследователей из австралийско-индонезийской команды, ведшей раскопки в Лианг-Буа, также совершили успешное плавание на бамбуковом плоту с Флореса на соседний остров Комодо, преодолев 22-километровый пролив с сильным течением[86]. Тем не менее в настоящее время Флорес — единственный исторически изолированный остров со следами пребывания гоминин миллион лет назад, что не поддерживает гипотезу о сознательных морских переходах. В то же время, возможно, что гоминины попали на остров случайно, на древесных стволах, унесённых тайфуном или цунами. Анализ местных течений показывает, что гоминины могли попасть на Флорес таким случайным образом с территории современных Камбоджи или Вьетнама через острова Калимантан и Сулавеси. Проблема с этой версией заключается в том, что древнейшим известным человеческим останкам на Сулавеси только 30 тысяч лет (хотя в 2016 году было сообщено о находке на острове каменных орудий возрастом свыше 100 тысяч лет, останки их создателей ещё не обнаружены[87]). Поэтому необходимо также учитывать возможность более прямого, хотя и более трудного маршрута с Явы (где в районе Сангирана обнаружены останки человека прямоходящего давностью 1,5 млн лет) через острова Бали, Ломбок и Сумбаву[61].
Среди сторонников гипотезы об отдельном виде не существует также единого мнения относительно того, когда этот вид окончательно вымер. Все известные находки были сделаны в слоях возрастом старше 17 тысяч лет[88]. На этом уровне в раскопах находится слой вулканического туфа, который археологи связывают с произошедшим в эти годы крупным вулканическим извержением[7] и выше которого, уже в слоях возрастом около 11,5 тысяч лет, найдены только костные останки, со всей очевидностью принадлежащие человеку разумному[47]. В 2009 году ван ден Берг и соавторы датировали время извержения и вымирания последних Homo floresiensis и карликовых стегодонов возрастом 17 тыс. лет назад[88]. Ещё одна гипотеза, связывающая исчезновение древних обитателей Флореса с современным человеком, относит его к более раннему периоду — от 50 до 40 тысяч лет назад, основываясь на датировке древнейших останков человека разумного на Флоресе[89]. С другой стороны, не исключено, что отдельные группы сумели пережить извержение и встречались в более поздние периоды с современными людьми, на что могут указывать местные легенды об эбу-гого — маленьких волосатых людях с покатыми лбами[90]. Согласно распространённой на Флоресе легенде, эбу-гого не только присутствовали на острове уже в исторические времена, но и совершали налёты на крестьянские хозяйства, за что и были истреблены полностью или частично[91]. Социальный антрополог Грегори Форт, ссылаясь на рассказы местных жителей, предполагает, что гоминиды, не относящиеся к виду Homo sapiens, могли выжить на Флоресе вплоть до XXI века[92].
Гипотезы о «больных хоббитах»
Из-за необычайно малого объёма черепной коробки LB1 у учёных сразу же возник вопрос, как же человек с такими размерами мозга мог изготовлять сложные орудия труда. Существование способных к изготовлению орудий разумных приматов с таким размером мозга, как у LB1, хотя и не противоречит в целом принципу островной карликовости, идёт вразрез с устоявшейся точкой зрения, согласно которой эволюционный прогресс сопряжён с увеличением соотношения массы мозга и тела, и с моделями уменьшения массы тела и мозга для островных карликовых видов. Для LB1 это соотношение слишком мало: разные модели предсказывают (если исходить из известных соотношений для Homo erectus) для такой массы мозга общую массу тела, не превышающую 11,6 килограмма, а в основном намного меньшую, между 2 и 6 килограммами[93][94] (известны, однако, исследования, как 2000-х годов, так и более ранние, показывающие, что для островных карликовых разновидностей животных стандартные модели соотношения размеров тела и мозга верны не всегда и бывают случаи, когда эти размеры уменьшаются синхронно[95][47]). Сомнения вызывает и способность стабильной популяции охотников-собирателей просуществовать на протяжении жизни тысяч поколений на таком маленьком острове, как Флорес[47].
Теория о разумном виде людей, развившемся по иной эволюционной линии, нежели человек разумный, и сосуществовавшем с ним, также противоречит гипотезе мультирегионального развития человека, согласно которой человек разумный представляет собой продукт независимой параллельной эволюции разных видов гоминин и которая не предполагает возможности альтернативной эволюции[27]. Именно теоретики-мультирегионалисты Мачей Хеннеберг и Алан Торн стали первыми, кто уже в 2004 году высказал предположение о том, что кости и каменные орудия с Флореса принадлежат не новому виду, а изуродованным патологией представителям Homo sapiens. Они предположили, что необычный вид Фло был результатом микроцефалии[96].

Теорию о микроцефалии подхватил и развил палеоантрополог Роберт Д. Мартин, а позже её поддержал и Теуку Якоб. Согласно сторонникам этой гипотезы, малый размер и форма мозга, укладывающиеся в рамки известных случаев микроцефалии[97], являются не единственным её доказательством. Другие симптомы этого заболевания могут включать собственно карликовость (проявляющуюся при как минимум одной разновидности микроцефалии — синдроме карликовости Маевского[англ.])[98], недоразвитую нижнюю челюсть со срезанным подбородком и слабыми зубами[99], слабые неразвитые мускулы и искривлённые кости[100]. Напротив, анатомические особенности, выделяемые сторонниками теории об отдельном виде (начиная с малого роста и включая отсутствие подбородка, приплюснутый нос и форму глазных орбит), как полагал Якоб, полностью укладываются в рамки внутрирасовых различий, характерных для представителей австраломеланезийской расы[101]. В 2007 году была высказана версия, согласно которой в условиях изоляции и инбридинга микроцефалия и ахондроплазия могли из случайного отклонения превратиться в определяющую характеристику и для целого племени[71].
В ответ на публикации сторонников гипотезы о микроцефалии, уже в 2007 году группа Дин Фальк опубликовала масштабное исследование, где эндокаст Фло сравнивался с многочисленными эндокастами известных современных микроцефалов, включая одного микроцефала-карлика. В статье были продемонстрированы существенные отличия между строением мозга LB1 и микроцефалов[102][103]. Тем не менее и четырьмя годами позже гипотеза о микроцефалии не считалась окончательно опровергнутой, как показывает анализ, проведённый Ваннуччи, Барроном и Холловеем. Эти авторы в очередной раз подчеркнули, что практически все особенности строения LB1 могут быть объяснены в рамках различных патологий, развивающихся при микроцефалии[104][105]. В начале 2016 года французские исследователи на основании микротомографических снимков черепа Фло признали наличие определённых патологий (в частности, фронтального внутреннего гиперостоза), но подчеркнули, что они не соответствуют симптомам, характерным для микроцефалии[33].
В 2006 году была выдвинута ещё одна гипотеза, объясняющая особенности строения LB1 в рамках известной патологии. Автор гипотезы, Гэри Ричардс, предположил, что Фло страдала от синдрома Ларона — генетического гормонального заболевания (это заболевание, в основном встречающееся у семитских народов, было примерно в это же время диагностировано и недалеко от Индонезии — на архипелаге Палау, где были обнаружены карликовые останки современных людей возрастом от 1400 до 2900 лет[77]). На следующий год группа исследователей, включавшая Цви Ларона, в честь которого названо этот синдром, выступила в поддержку теории Ричардса, подчеркивая, что симптомы заболевания практически неотличимы от наличествующих у Фло[106]. Синдром Ларона характерен в основном для семитских народов Ближнего Востока, но не ограничивается ими[107]. Критики этой теории, в первую очередь Дин Фальк, отмечают, что несмотря на сходство описаний, общий физический вид больных синдромом Ларона значительно отличается от предполагаемого облика человека флоресского. Более того, отмечается, что новый список симптомов значительно отличается от традиционного, включавшего тонкие кости и череп. Эти характеристики, а также неразвитые лобные пазухи резко отличают традиционные симптомы синдрома Ларона от относительно толстокостной Фло, обладавшей мощными лобными пазухами[108].
Более новая гипотеза, сформулированная в 2007 году, связывает облик обитателей пещеры Лианг-Буа с микседематозным кретинизмом (следствие тяжелого йододефицита у матери при вынашивании ребёнка). Учёные отмечают, что характерными признаками данного заболевания являются тяжёлая олигофрения, замедление физического развития, короткие конечности, большая голова с широким уплощённым носом, а рост при этом составляет не более одного метра. Авторы теории дискутируют со сторонниками гипотезы, связанной с синдромом Ларона, подчёркивая тонкие черты, длинные ступни, развитые подбородки и выпуклые лбы пациентов, страдающих от этой болезни, по сравнению с более грубым обликом, характерным как для кретинов, так и для LB1; они также отмечают, что на островах Индонезии заболевания, связанные с недостатком йода, давно известны и широко распространены, а в последние годы обнаружены и случаи недостатка селена[109]. В следующие несколько лет были опубликованы как мнения в поддержку этой гипотезы (опирающиеся на факт широкого диапазона характеристик людей, страдающих различными формами кретинизма[110], среди которых, как и среди характеристик различных форм микроцефалии, можно подыскать соответствующие описанию флоресских скелетов), так и её опровержения[111]; Питер Браун отмечает, что кроме малого роста у останков из Лианг-Буа и современных кретинов только одна общая черта — высокий угол поворота плечевой кости, но эту же черту разделяют также останки Homo erectus из Дманиси (возраст которых приближается к 2 млн лет)[112].
Согласно ещё одной альтернативной гипотезе, обнародованной в 2008 году, малый рост и некоторые примитивные черты LB1 могут быть результатом первичной карликовости (разновидность, при которой отставание в росте начинается ещё на стадии беременности), которая в свою очередь является следствием мутаций гена PCNT, расположенного на 21-й хромосоме[113].
В 2014 году Мачей Хеннеберг с соавторами выдвинул новую гипотезу, согласно которой физические характеристики Фло указывают на то, что она страдала от синдрома Дауна. В статье утверждается, что антропометрические данные Фло, включая лицевую асимметрию (проявляющуюся в неравномерном износе зубов) и укороченную бедренную кость, полностью укладываются в диагностическую картину синдрома Дауна. Авторы гипотезы считают, что с поправкой на это заболевание рост Фло не является исключительным для представителей современных австраломеланезийских народов. Проблемы с работой щитовидной железы наблюдаются, как указывает Хеннеберг, более чем у трети пациентов с синдромом Дауна, и предположение о том, что Фло страдала одновременно от кретинизма и синдрома Дауна, по мнению авторов полностью объясняет все её физические особенности[114].
Сразу же после публикации новой гипотезы её авторам были предъявлены обвинения в методологической недобросовестности и возможных нарушениях научной этики. Журнал PNAS, в котором публиковались все новые теории группы Хеннеберга, был обвинён в пренебрежении обычной процедурой научного рецензирования, так как он в целях ускорения публикаций предоставляет членам Национальной академии наук США право самим выбирать себе рецензентов. В данном случае такое право получил постоянный соавтор теорий Хеннеберга, американский геолог Кеннет Сюй. Рассматривая методологию новой публикации, Дин Фальк подчеркнула, что в статье не приводится ни одной сравнительной фотографии скелетов больных синдромом Дауна и человека флоресского, что сразу показало бы несостоятельность нового сравнения, а австралиец Уильям Юнгерс задался вопросом, известны ли среди больных синдромом Дауна индивиды с черепной коробкой объёмом 400 кубических сантиметров[115]. Вскоре после выхода статьи Хеннеберга в журнале PNAS было опубликовано письмо, авторы которого — антропологи и археологи из трёх стран — пишут об отличиях устройства двух известных нижних челюстей из Лианг-Буа (LB1 и LB6) от челюстей современного человека, называя несостоятельным одно из ключевых утверждений Хеннеберга — о частом проявлении у современных австраломеланезийцев такой архаичной черты, как отсутствие подбородочного выступа, а также указывая на наличие у челюстей с Флореса внутреннего утолщения подбородочного шва[англ.], характерного для древних видов рода Homo, но не для Homo sapiens[116]. Браун, Фальк, Юнгерс и другие соавторы опубликованного в 2016 году исследования, сравнивающего фенотипические характеристики LB1 и современных пациентов с синдромом Дауна, пришли к выводу, что имеющегося сходства недостаточно для вывода о том, что LB1 страдала от этого заболевания, и что её черты по-прежнему можно считать уникальными[117].
Среди прочих аргументов против теории об отдельном виде может быть упомянут курьёз, связанный с утверждением, что первый левый моляр в нижней челюсти LB1 содержит пломбу, типичную для стоматологов, работавших в Индонезии в 30-е годы XX века. Если бы это утверждение, которое в 2008 году сделал достоянием гласности Хеннеберг, оказалось верным, история Homo floresiensis на этом была бы окончена. Однако утверждение было сначала опровергнуто Питером Брауном, который лично исследовал каждый зуб[118], а затем и отдельным исследованием, опубликованным в 2011 году[119]. По-видимому, за материал пломбы был принят побелевший в известняковой пещере дентин, открывшийся под стёртой эмалью[118].
В целом Хеннеберг, Роберт Экхардт из Университета штата Пенсильвания и другие противники теории о видовой самостоятельности Homo floresiensis строят свою позицию на том факте, что за годы раскопок, прошедшие после объявления об открытии нового вида людей, не было найдено ни одного скелета, обладающего теми же анатомическими особенностями, что и Фло, ставшая его типовым представителем. С их точки зрения, это доказывает, что никакого отдельного вида не существует, а найденные в Лианг-Буа останки принадлежат представителям местной низкорослой расы человека разумного[38]. Критики теории отдельного вида указывают, что любая из специфичных черт LB1 либо укладывается в диапазон нормальных характеристик Homo sapiens (как, например, очень малый рост), либо может быть объяснена патологическими изменениями организма. Согласно сторонникам этой позиции, сочетание патологически маленькой черепной коробки, низкого роста и специфического соотношения костей конечностей не является статистически невозможным, а значит, в соответствии с принципом бритвы Оккама, не следует изобретать новые сущности. Вне зависимости от того, могут ли они предложить непротиворечивый диагноз патологий, характеризующих Фло, они настаивают, что сам факт патологических изменений организма невозможно отрицать и он опровергает мнение, что эти останки могут служить голотипом нового вида[120].
Другие версии
Хотя основное противостояние в вопросе классификации останков из пещеры Лианг-Буа проходит между сторонниками гипотезы об отдельном виде и сторонниками гипотезы о патологически изменённом представителе Homo sapiens, существуют также точки зрения, отличные от обеих господствующих гипотез. Так, в 2013 году была выдвинута версия, согласно которой флоресские останки могут принадлежать целому карликовому народу современных людей, для которых малый рост и объём черепа — не патология. Авторы статьи ссылаются на найденные в 2008 году в Палау карликовые останки, возраст которых составляет от 900 до 2900 лет. Эти останки несомненно принадлежат людям современного типа, но при этом помимо малого роста — от 0,94 до 1,2 м — обладают небольшим (хотя и всё ещё вдвое бо́льшим, чем у LB1) объёмом черепа и некоторыми примитивными чертами. Таким образом, предполагается, что островная карликовость нормальна и для современных людей, что ранее отрицалось[72].
Другой радикальный взгляд на флоресские останки был озвучен одним из ранних сторонников версии о микроцефалии[121] — Альфредом Чарнецки. Согласно его версии, кости из Лианг-Буа принадлежат орангутанам[122]. Однако с этой гипотезой не согласились даже бывшие единомышленники Чарнецки — Экхардт и Хеннеберг, указавшие, что его выводы построены на неполной информации в условиях отсутствия доступа к останкам и не учитывают описанные в последние годы черты LB1, не присущие орангутанам[123].
Примечания
- ↑ 1 2 Sutikna, T., et al. Revised stratigraphy and chronology for Homo floresiensis at Liang Bua in Indonesia (англ.) // Nature. — 30 March 2016. — doi:10.1038/nature17179.
- ↑ Morwood & van Oosterzee, 2007, pp. 48—51.
- ↑ Dalton, R. Little lady of Flores forces rethink of human evolution (англ.) // Nature. — 28 October 2004. — Vol. 431. — P. 1029. — doi:10.1038/4311029a. Архивировано 4 октября 2011 года.
- ↑ 1 2 3 Ivereigh, D. Bones of contention (англ.). Cosmos (октябрь 2007). Дата обращения: 14 сентября 2012. Архивировано из оригинала 7 февраля 2016 года.
- ↑ Morwood & van Oosterzee, 2007, p. 34.
- ↑ Morwood & van Oosterzee, 2007, pp. 86—88.
- ↑ 1 2 Sarmiento et al., 2007, p. 178.
- ↑ van den Bergh, G. D., Meijer, H. J. M., Rokus Awe Due et al. The Liang Bua faunal remains: a 95 k.yr. sequence from Flores, East Indonesia (англ.) // Journal of Human Evolution. — 2009. — Vol. 57, no. 5. — P. 527. — doi:10.1016/j.jhevol.2008.08.015.
- ↑ 1 2 3 4 Morwood, M. J., Brown, P., Jatmiko et al. Further evidence for small-bodied hominins from the Late Pleistocene of Flores, Indonesia (англ.) // Nature. — 13 October 2005. — Vol. 437. — P. 1012—1017. — doi:10.1038/nature04022. Архивировано 5 марта 2016 года.
- ↑ Cauchi, S. Lost race of human 'hobbits' (англ.). The Age (28 октября 2004). Дата обращения: 13 сентября 2012. Архивировано 23 октября 2012 года.
- ↑ Morwood & van Oosterzee, 2007, pp. 100, 110, 151—153.
- ↑ 1 2 3 Aiello, 2010, p. 169.
- ↑ Morwood & van Oosterzee, 2007, p. 95.
- ↑ Morwood & van Oosterzee, 2007, pp. 188—194.
- ↑ Morwood & van Oosterzee, 2007, pp. 88—89.
- ↑ 1 2 Hobbits triumph tempered by tragedy (англ.). Sydney Morning Herald (5 марта 2005). Дата обращения: 15 сентября 2012. Архивировано 23 октября 2012 года.
- ↑ Falk, 2011, pp. 92—95.
- ↑ 1 2 Hobbit cave digs set to restart (англ.). BBC News (25 января 2007). Дата обращения: 15 сентября 2012. Архивировано 23 октября 2012 года.
- ↑ Powledge, T. Flores hominid bones returned (англ.). The Scientist (28 февраля 2005). Дата обращения: 15 сентября 2012. Архивировано 23 октября 2012 года.
- ↑ Morwood & van Oosterzee, 2007, p. 233.
- ↑ Gagan, M. K., et al. Geoarchaeological finds below Liang Bua (Flores, Indonesia): A split-level cave system for Homo floresiensis? // Palaeogeography, Palaeoclimatology, Palaeoecology. — 2015. — Vol. 440. — P. 533—550. — doi:10.1016/j.palaeo.2015.09.021.
Barras, C. Frodo’s basement: Secret chamber found where hobbit humans lived. New Scientist (29 октября 2015). Дата обращения: 2 ноября 2015. Архивировано 31 октября 2015 года. - ↑ 1 2 3 Alex B. 'The hobbit' may have shrunk early, evolved from a tall human ancestor (англ.) // Science. — 2024. — 6 August. — doi:10.1126/science.zhidvfr. Архивировано 7 августа 2024 года.
- ↑ Argue, D., Donlon, D., Groves, C. and Wright, R. Homo floresiensis: Microcephalic, pygmoid, Australopithecus, or Homo? (англ.) // Journal of Human Evolution. — Elsevier, 2006. — Vol. 51. — P. 360—374. — doi:10.1016/j.jhevol.2006.04.013. Архивировано 22 февраля 2016 года.
- ↑ 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 Jungers, 2012.
- ↑ Baten, J. Global Height Trends in Industrial and Developing Countries, 1810-1984: An Overview // Working Paper. — University of Tubingen, 2006.
- ↑ Weber, G. The Andamanese: A Physical Examination (англ.). George Weber's Lonely Islands: The Negrito People and the Out-of-Africa Story of the human race (3 августа 2005). Дата обращения: 14 сентября 2012. Архивировано из оригинала 10 июля 2012 года.
- ↑ 1 2 Lahr, M. M. and Foley, R. Human evolution writ small (англ.) // Nature. — 28 October 2004. — Vol. 431. — P. 1043—1044. — doi:10.1038/4311029a. Архивировано 31 декабря 2019 года.
- ↑ 1 2 Weston, E. M. and Lister, A. M. Insular dwarfism in hippos and a model for brain size reduction in Homo floresiensis (англ.) // Nature. — 2009. — Vol. 459. — P. 85—88. — doi:10.1038/nature07922. Архивировано 27 января 2022 года.
- ↑ Morwood & van Oosterzee, 2007, p. 86.
- ↑ Falk, 2011, p. 138.
- ↑ 1 2 Vanucci, R. C., Barron, T. F. and Holloway, R. L. Craniometric ratios of microcephaly and LB1, Homo floresiensis, using MRI and endocasts // Proceedings of the National Academy of Sciences. — 2011. — Vol. 108, № 34. — P. 14043—14048. — doi:10.1073/pnas.1105585108. Архивировано 7 ноября 2017 года.
- ↑ 1 2 Gordon, A. D., Nevell, L. and Wood, B. The Homo floresiensis cranium (LB1): Size, scaling, and early Homo affinities // Proceedings of the National Academy of Sciences. — 2008. — Vol. 105, № 12. — P. 4650—4655. — doi:10.1073/pnas.0710041105. Архивировано 24 сентября 2015 года.
- ↑ 1 2 Balzeau, A., and Charlier, P. Les origines d'Homo floresiensis d'après l'intérieur de son crâne (фр.). Hominidés.com (15 февраля 2016). Дата обращения: 16 февраля 2016. Архивировано 25 марта 2016 года.
- ↑ Falk, 2011, pp. 116—118.
- ↑ Falk, 2011, p. 119.
- ↑ Falk, 2011, p. 118.
- ↑ Falk, 2011, p. 120—122.
- ↑ 1 2 Eckhardt, R. B., Henneberg, M., Chavanaves, S., Weller, A. S. and Hsü, K. J.. Reply to Westaway et al.: Mandibular misrepresentations fail to support the invalid species Homo floresiensis // Proceedings of the National Academy of Sciences. — 2015. — Vol. 112, № 7. — P. E606. — doi:10.1073/pnas.1422176112. Архивировано 20 февраля 2015 года.
- ↑ Brown, P. and Maeda, T. Liang Bua Homo floresiensis mandibles and mandibular teeth: a contribution to the comparative morphology of a new hominin species // Journal of Human Evolution. — 2009. — Vol. 57. — P. 571—596. — doi:10.1016/j.jhevol.2009.06.002.
- ↑ Kaifu, Y., Kono, R. T., Sutikna, T., Saptomo, E. W., Jatmiko, et al. Unique dental morphology of Homo floresiensis and its evolutionary implications // PLoS ONE. — 2015. — Vol. 10, № 11. — doi:10.1371/journal.pone.0141614. Архивировано 1 февраля 2016 года.
Человек флоресский, aka «хоббит», не был одного вида с современным человеком. Научная Россия (23 ноября 2015). Дата обращения: 19 января 2016. Архивировано 26 января 2016 года. - ↑ 1 2 3 Kaifu Y., et al. Early evolution of small body size in Homo floresiensis (англ.) // Nature Communications. — 2024. — Vol. 15, no. 6381. — doi:10.1038/s41467-024-50649-7. Архивировано 11 августа 2024 года.
- ↑ Jacob, T., Indriati, E., Soejono, R. P., Hsu, K., Frayer, D. W., Eckhardt, R. B., Kuperavage, A. J., Thorne, A. and Henneberg, M. Pygmoid Australomelanesian Homo sapiens skeletal remains from Liang Bua, Flores: Population affinities and pathological abnormalities // Proceedings of the National Academy of Sciences. — 2008. — Vol. 103, № 36. — P. 13421—13426. — doi:10.1073/pnas.0605563103.
- ↑ Larson, S. G., Jungers, W. L., Morwood, M. J., Sutikna, T., Jatmiko, Saptomo, E. W., Rokus Awe Due and Djubiantono, T. Homo floresiensis and the evolution of the hominin shoulder (англ.) // Journal of Human Evolution. — Elsevier, 28 October 2004. — Vol. 53, no. 6. — P. 718—731. — doi:10.1016/j.jhevol.2007.06.003. Архивировано 13 июня 2011 года.
- ↑ 1 2 Марков А. Примитивное строение запястья подтверждает принадлежность хоббитов к особому виду людей. Элементы.ру (24 сентября 2007). Дата обращения: 14 сентября 2012. Архивировано 23 октября 2012 года.
- ↑ Salleh, A. Wrist gives hobbit theory the flick (англ.). ABC.net.au (21 сентября 2007). Дата обращения: 14 сентября 2012. Архивировано 23 октября 2012 года.
- ↑ Flohr S. Did the assumed partial skeleton LB1 (aka Homo floresiensis) really have long feet? (англ.) // Anthropologischer Anzeiger; Bericht uber die biologisch-anthropologische Literatur. — 2018. — Vol. 75, no. 2. — P. 169—174. — doi:10.1127/anthranz/2018/0770.
- ↑ 1 2 3 4 Марков, 2012.
- ↑ Jungers, W. L., Larson, S. G., Harcourt-Smith, W., Morwood, M. J., Sutikna, T., Awe, R. D., and Djubiantono, T. Descriptions of the lower limb skeleton of Homo floresiensis // Journal of Human Evolution. — 2009. — Vol. 57. — P. 538—554. — doi:10.1016/j.jhevol.2008.08.014. Архивировано 15 марта 2016 года.
- ↑ Argue et al., 2006, pp. 370—371.
- ↑ Bezzi, L. Homo floresiensis. ATOR (Arc-Team Open Research) (13 августа 2015). Дата обращения: 22 января 2016. Архивировано 28 января 2016 года.
- ↑ Falk, 2011, p. 78.
- ↑ Thompson, H. Paleoartist Brings Human Evolution to Life (англ.). Smithsonian.com (17 мая 2014). Дата обращения: 21 января 2016. Архивировано 27 января 2016 года.
Urban, M. La dame de Florès au musée de l'Homme (фр.). RFI (11 декабря 2007). Дата обращения: 21 января 2016. Архивировано 31 января 2016 года. - ↑ Hall, S. UOW hobbit makes it big in Japan (англ.). Illawarra Mercury (26 апреля 2010). Дата обращения: 21 января 2016. Архивировано 26 января 2016 года.
- ↑ Hayes, S., Sutikna, T., and Morwood, M. Faces of Homo floresiensis (LB1) // Journal of Archaeological Science. — 2013. — Vol. 40, № 12. — P. 4400—4410. Архивировано 27 января 2016 года.
- ↑ UOW plays host to Australian Archaeological Conference (англ.). University of Wollongong (10 декабря 2012). Дата обращения: 12 декабря 2012. Архивировано 16 декабря 2012 года.
- ↑ Wong, K. Reconstructed Face of Extinct “Hobbit” Species Is Startlingly Humanlike (англ.). Scientific American Blog (11 декабря 2012). Дата обращения: 12 декабря 2012. Архивировано 16 декабря 2012 года.
- ↑ Маркина Н. Хоббит не патология, а вид человека. Газета.Ru (15 июля 2013). Дата обращения: 15 июля 2013. Архивировано 4 сентября 2014 года.
Karen L. Baab, Kieran P. McNulty, Katerina Harvati[англ.]. Homo floresiensis Contextualized: A Geometric Morphometric Comparative Analysis of Fossil and Pathological Human Samples // PLOS ONE. — 2013. — Vol. 8(7). — doi:10.1371/journal.pone.0069119. Архивировано 4 сентября 2014 года. - ↑ Dynamic similarity predicts gait parameters for Homo floresiensis and the Laetoli hominins // American Journal of Human Biology. — 2008. — Vol. 20, № 3. — P. 312—316. — doi:10.1002/ajhb.20721. Архивировано 9 апреля 2016 года.
- ↑ Flores 'hobbit' walked more like a clown than Frodo (англ.). New Scientist (16 апреля 2008). Дата обращения: 21 января 2016. Архивировано 25 марта 2016 года.
- ↑ Brumm, A., et al. Hominins on Flores, Indonesia, by one million years ago // Nature. — 2010. — Vol. 464, № 7289. — P. 748—752. — doi:10.1038/nature08844.
- ↑ 1 2 3 Dennell, R. W., Louys, J., O'Regan, H. J. and Wilkinson, D. M. The origins and persistence of Homo floresiensis on Flores: biogeographical and ecological perspectives // Quaternary Science Reviews. — 2014. — Vol. 96. — P. 98—107. — doi:10.1016/j.quascirev.2013.06.031.
- ↑ Moore, M. W. and Brumm, A. Homo floresiensis and the African Oldowan // Interdisciplinary approaches to the Oldowan / Erella Hovers, David R. Braun (Eds.). — Dordrecht: Springer, 2009. — P. 61—69. — ISBN 978-1-4020-9060-8.
- ↑ Moore, M. W., Sutikna, T., Morwood, M. J. and Brumm, A. Continuities in stone flaking technology at Liang Bua, Flores, Indonesia // Journal of Human Evolution. — 2009. — Vol. 57, № 5. — P. 503—526. — doi:10.1016/j.jhevol.2008.10.006.
- ↑ Culotta, E. Did Humans Learn From Hobbits? // Science. — 2009. — Vol. 324, № 5926. — P. 447. — doi:10.1126/science.324_447.
- ↑ Aiello, 2010, p. 176.
- ↑ Falk, 2011, p. 88.
- ↑ Fire discovery sheds new light on 'hobbit' demise. Science Daily (29 июня 2016). Дата обращения: 10 ноября 2016. Архивировано 11 ноября 2016 года.
- ↑ Falk, 2011, pp. 88—90.
- ↑ Falk, 2011, p. 134.
- ↑ Jones, C. Researchers to drill for hobbit history. Nature (5 января 2011). Дата обращения: 20 января 2016. Архивировано 23 января 2016 года.
- ↑ 1 2 Linden, A. Reclassification of Homo floresiensis as Homo species indeterminate to accurately reflect a lack of existing knowledge // Totem: The University of Western Ontario Journal of Anthropology. — 2007. — Vol. 15, № 1. — P. 5—20. Архивировано 29 июля 2013 года.
- ↑ 1 2 De Cruz, H. and De Smedt, J. The value of epistemic disagreement in scientific practice. The case of Homo floresiensis // Studies in History and Philosophy of Science. Part A. — 2013. — Vol. 44, № 2. — P. 169—177. — doi:10.1016/j.shpsa.2013.02.002. Архивировано 6 марта 2016 года.
- ↑ 1 2 3 Linden, 2007, p. 8.
- ↑ Bromham, L. and Cardillo, M. Primates follow the ‘island rule’: implications for interpreting Homo floresiensis // Biology Letters: Evolutionary Biology. — 2007. — Vol. 3. — P. 398—400. — doi:10.1098/rsbl.2007.0113. Архивировано 31 августа 2021 года.
- ↑ Morwood & van Oosterzee, 2007, pp. 177—178.
- ↑ 1 2 3 4 5 Argue, D., Morwood, M. J., Sutikna, T., Jatmiko and Saptomo, E. W. Homo floresiensis: a cladistic analysis // Journal of Human Evolution. — 2009. — Vol. 57, № 5. — P. 623—639. — doi:10.1016/j.jhevol.2009.05.002.
- ↑ 1 2 Bednarik, R. G. Reviewing the Flores Hobbit Chronicles // Anthropos. — 2009. — Bd. 104. — S. 195—198. Архивировано 17 марта 2016 года.
- ↑ Villmoare, B. Early Homo and the role of the genus in paleoanthropology (англ.) // American Journal of Physical Anthropology. — 2018. — Vol. 165. — P. 80. — doi:10.1002/ajpa.23387.
- ↑ Strait D. S., and Grine F. Inferring hominoid and early hominid phylogeny using craniodental characters: The role of fossil taxa (англ.) // Journal of Human Evolution. — Elsevier, 2004. — Vol. 47, no. 6. — P. 399—452. — doi:10.1016/j.jhevol.2004.08.008.
- ↑ 1 2 Dembo, M., Matzke, N. J., Mooers, A. Ø. and Collard, M. Bayesian analysis of a morphological supermatrix sheds light on controversial fossil hominin relationships // Proceedings of the Royal Society B: Biological Sciences. — 2015. — Vol. 282, № 1812. — P. 20150943. — doi:10.1098/rspb.2015.0943. Архивировано 27 ноября 2018 года.
- ↑ Марков А. Предками хоббитов могли быть люди из Дманиси. Элементы.ру (7 мая 2009). Дата обращения: 26 июля 2015. Архивировано 25 марта 2016 года.
- ↑ Falk, 2011, p. 184.
- ↑ Argue, D., Groves, C. P., Lee, M. S. and Jungers, W. L. The affinities of Homo floresiensis based on phylogenetic analyses of cranial, dental, and postcranial characters // Journal of Human Evolution. — 2017. — Vol. 30. — P. 1—27. — doi:10.1016/j.jhevol.2017.02.006. Архивировано 25 апреля 2017 года.
Origins of Indonesian Hobbits finally revealed. Science Daily (21 апреля 2017). Дата обращения: 26 апреля 2017. Архивировано 24 апреля 2017 года. - ↑ van den Bergh, G. D., et al. Homo floresiensis-like fossils from the early Middle Pleistocene of Flores // Nature. — 2016. — Vol. 534. — P. 245—248. — doi:10.1038/nature17999. Архивировано 12 февраля 2021 года.
Adam Hoffman. New Fossils Hint 'Hobbit' Humans Are Older Than Thought. National Geographic (8 июня 2016). Дата обращения: 8 июня 2016. Архивировано 9 июня 2016 года. - ↑ Sarmiento et al., 2007, p. 177.
- ↑ Morwood & van Oosterzee, 2007, pp. 197—200.
- ↑ Антонов Е. Первые люди в Индонезии появились раньше, чем считалось. Наука и жизнь (19 января 2016). Дата обращения: 30 марта 2016. Архивировано 12 апреля 2016 года.
- ↑ 1 2 van den Bergh et al., 2009, p. 528.
- ↑ Callaway, E. Human remains found in hobbit cave. Nature (21 сентября 2016). Дата обращения: 10 ноября 2016. Архивировано 25 сентября 2016 года.
- ↑ Морвуд, М., Сутикна, Т. и Робертс, Р. Находка на острове Флорес. National Geographic-Россия (апрель 2005). Дата обращения: 16 сентября 2012. Архивировано 16 июля 2019 года.
Мужчинкин, В. «Хоббиты» с острова Цветов // Наука и жизнь. — 2005. — № 11. Архивировано 14 августа 2014 года. - ↑ Redfern, N. The Bigfoot Book: The Encyclopedia of Sasquatch, Yeti and Cryptid Primates. — Canton,MI: Visible Ink, 2016. — P. 109, 120—121. — ISBN 978-1-57859-561-7. Архивировано 30 января 2016 года.
- ↑ Ly L. Everything old is new again (англ.). Work of Arts (19 февраля 2015). Архивировано 16 августа 2018 года.
Forth G. Opinion: Another Species of Hominin May Still Be Alive (англ.). The Scientist (18 апреля 2022). Дата обращения: 19 апреля 2022. Архивировано 19 апреля 2022 года. - ↑ Martin et al. (Anatomical Records), 2006, pp. 1126—1127.
- ↑ Martin, R. D., MacLarnon, A. M., Phillips, J. L., Dussubieux, L., Williams, P. R. and Dobyns, W. B. Comment on "The Brain of LB1, Homo floresiensis" (англ.) // Science. — 2006. — Vol. 312. — P. 999. — doi:10.1126/science.1121144.
- ↑ Falk, 2011, pp. 124—125.
- ↑ Falk, 2011, pp. III—IV.
- ↑ Martin et al. (Anatomical Records), 2006.
- ↑ Martin et al. (Anatomical Records), 2006, p. 1136.
- ↑ Martin et al. (Anatomical Records), 2006, p. 1140.
- ↑ Jacob et al., 2008, pp. 13424—13425.
- ↑ Jacob et al., 2008, pp. 13422—13424.
- ↑ Falk, D., Hildebolt, C., Smith, K., et al. Brain shape in human microcephalics and Homo floresiensis // Proceedings of the National Academy of Sciences. — 2007. — Vol. 104, № 7. — P. 2513—2518. — doi:10.1073/pnas.0609185104. Архивировано 1 октября 2007 года.
- ↑ Falk, 2011, pp. 142—147.
- ↑ Vanucci, Barron and Holloway, 2011.
- ↑ Соколов, А. Homo floresiensis: 3:2 в пользу микроцефалии… Антропогенез.ру (25 августа 2011). Дата обращения: 17 сентября 2012. Архивировано 23 октября 2012 года.
- ↑ Hershkovitz, I., Kornreich, L. and Laron, Z. Comparative skeletal features between Homo floresiensis and patients with primary growth hormone insensitivity (Laron syndrome) // American Journal of Physical Anthropology. — 2007. — Vol. 134, № 2. — P. 198—208. — doi:10.1002/ajpa.20655. Архивировано 11 января 2012 года.
- ↑ Galli-Tsinopoulou, A., Nousia-Arvanitakis, S., Tsinopoulos, I., Bechlivanides, C., Shevah, O. and Laron, Z. Laron syndrome. First report from Greece // Hormones. — 2003. — Vol. 2, № 2. — P. 120—124. Архивировано 13 декабря 2010 года.
Rosenbloom, A. L. Growth Hormone Insensitivity Syndrome // Pediatric Endocrinology: A Practical Clinical Guide / S. Radovick, M. H. MacGillivray (Eds.). — Humana Press, 2003. — P. 43. — ISBN 978-0-89603-946-9. - ↑ Falk, 2011, pp. 152155.
- ↑ Obendorf, P. J., Oxnard, C. E. and Kefford, B. J. Are the small human-like fossils found on Flores human endemic cretins? // Proceedings of the Royal Society B. — 2007. — Vol. 275, № 1640. — P. 1287—1296. — doi:10.1098/rspb.2007.1488.
- ↑ Oxnard, C. E., Obendorf, P. J. and Kefford, B. J. Post-Cranial Skeletons of Hypothyroid Cretins Show a Similar Anatomical Mosaic as Homo floresiensis // PLOS ONE. — 2010. — Vol. 5, № 10. — doi:10.1371/journal.pone.0013018. Архивировано 28 сентября 2012 года.
- ↑ Falk, 2011, pp. 156—158.
- ↑ Brown, P. LB1 and LB6 Homo floresiensis are not modern human (Homo sapiens) cretins (англ.) // Journal of Human Evolution. — Elsevier, 2012. — Vol. 62, no. 2. — P. 201—224. — doi:10.1016/j.jhevol.2011.10.011. Архивировано 24 сентября 2015 года.
- ↑ Rauch, A. et al. Mutations in the Pericentrin (PCNT) Gene Cause Primordial Dwarfism (англ.) // Science. — 2008. — Vol. 319, no. 5864. — P. 816—819. — doi:10.1126/science.1151174.
Kamrani, K. Is the Homo floresiensis phenotype due to mutations in the PCNT gene? Anthropology.net (8 января 2008). Дата обращения: 16 октября 2015. Архивировано из оригинала 9 сентября 2019 года. - ↑ Henneberg, M., Eckhardt, R. B., Chavanaves, S. and Hsü, K. J. Evolved developmental homeostasis disturbed in LB1 from Flores, Indonesia, denotes Down syndrome and not diagnostic traits of the invalid species Homo floresiensis // Proceedings of the National Academy of Sciences. — 2014. — Vol. 111, № 33. — P. 11967—11972. — doi:10.1073/pnas.1407382111. Архивировано 13 марта 2015 года.
Nogrady, B. Did the 'Hobbit' have Down syndrome? ABC Science (5 августа 2014). Дата обращения: 29 августа 2014. Архивировано 31 августа 2014 года. - ↑ McKie, R. Homo floresiensis: scientists clash over claims 'hobbit man' was modern human with Down's syndrome. The Guardian (17 августа 2014). Дата обращения: 10 сентября 2014. Архивировано 25 марта 2016 года.
- ↑ Westaway, M. C., Durband, A. C., Groves, C. P. and Collard M. Mandibular evidence supports Homo floresiensis as a distinct species // Proceedings of the National Academy of Sciences. — 2015. — Vol. 112, № 7. — P. E604—E605. — doi:10.1073/pnas.1418997112. Архивировано 20 февраля 2015 года.
- ↑ Baab, K., et al. A Critical Evaluation of the Down Syndrome Diagnosis for LB1, Type Specimen of Homo floresiensis // PLOS One. — 2016. — Vol. 11, № 6. — doi:10.1371/journal.pone.0155731.
- ↑ 1 2 Falk, 2011, p. 158—160.
- ↑ Jungers, W. L. and Kaifu, Y. On dental wear, dental work, and oral health in the type specimen (LB1) of Homo floresiensis // American Journal of Physical Anthropology. — 2011. — Vol. 145, № 2. — P. 282—289. — doi:10.1002/ajpa.21492. Архивировано 26 ноября 2015 года.
- ↑ Eckhardt, R. B., Henneberg, M., Weller, A. S. and Hsü K. J. Rare events in earth history include the LB1 human skeleton from Flores, Indonesia, as a developmental singularity, not a unique taxon // Proceedings of the National Academy of Sciences. — 2014. — Vol. 111, № 33. — P. 11961—11966. — doi:10.1073/pnas.1407385111. Архивировано 5 декабря 2015 года.
- ↑ Weber, J., Czarnetzki, A. and Pusch, C. M. Comment on "The Brain of LB1, Homo floresiensis" // Science. — 2005. — Vol. 310, № 5746. — P. 236b.
- ↑ Czarnetzki, A. The unravelled LB1 (Homo floresiensis) riddle? Some critical comments on the morphology of LB1 // Archäologische Informationen. — 2015. — Vol. 37. — P. 181—189. Архивировано 11 июля 2015 года.
- ↑ Eckhardt, R. B., Chavanaves, S. and Henneberg, M. Riddle Unravel Reprised: the role of LB1 in inferences about human functional morphology and phylogeny // Archäologische Informationen. — 2015. — Vol. 38.
Литература
- Aiello, L. C. Five Years of Homo floresiensis (англ.) // American Journal of Physical Anthropology. — Wiley, 2010. — Vol. 142, no. 2. — P. 167—179. — doi:10.1002/ajpa.21255. Архивировано 7 июля 2014 года.
- Falk, D. The Fossil Chronicles: How Two Controversial Discoveries Changed Our View of Human Evolution (англ.). — University of California Press, 2011. — ISBN 978-0-520-26670-4.
- Henneberg, M., and Schofield, J. The Hobbit Trap: Money, Fame, Science and the Discovery of a 'new Species' (англ.). — Kent Town, Australia: Wakefield Press, 2008. — ISBN 978-1-86254-791-9.
- Jungers, W. L. Homo floresiensis // A Companion to Paleoanthropology (англ.) / David R. Begun (Ed.). — John Wiley and Sons, 2012. — (Blackwell Companions to Antropology). — ISBN 9781118332375.
- Martin, R. D., MacLarnon, A. M., Phillips, J. L. and Dobyns, W. B. Flores Hominid: New Species or Microcephalic Dwarf? (англ.) // The Anatomical Records, Part A. — 2006. — Vol. 288A. — P. 1123—1145. — doi:10.1002/ar.a.20389.
- Morwood, M., and van Oosterzee, P. A New Human (англ.). — New York: Smithsonian Books, 2007. — ISBN 978-0-06-089908-0.
- Sarmiento, Esteban E., Sawyer, Gary J. , and Milner, Richard. An island sunset for the little people: Homo floresiensis // The Last Human: A Guide to Twenty-two Species of Extinct Humans (англ.). — Yale University Press, 2007. — P. 176—181. — ISBN 978-0-300-10047-1.
- Марков Александр. «Хоббиты» с острова Флорес // Эволюция человека. — М.: Астрель-Corpus, 2012. — Т. 1. Обезьяны, кости и гены. — С. 182—200. — ISBN 978-5-271-36293-4.
| Таксономия | |
|---|---|
| В библиографических каталогах |
| Вымершие роды Гоминини/Хоминина | |
|---|---|
| Люди (род Homo) | |
| Находки гоминид | |
| Происхождение |
|
| Распространение | |
Вымершие приматы | |||||||
|---|---|---|---|---|---|---|---|
| Базальные таксоны | |||||||
| Вымершие полуобезьяны |
| ||||||
| Вымершие обезьяны | |||||||
| Гоминиды | |||||||