Череп птиц
Череп птиц — часть скелета птиц. Отличается большой черепной коробкой, бесшовным соединением многих костей и клювом, который заменил у них челюсти с зубами. Челюстной аппарат птиц, их клюв и язык у разных видов очень отличаются формой и размером. Это одна из самых вариабельных систем органов птиц.
Способность верхней челюсти перемещаться вертикально относительно черепной коробки называется кинетичностью черепа. Если гибкая зона лежит у основания надклювья, то говорят о прокинетизме, а если она расположена на уровне носовых ям и верхняя челюсть обладает внутренней подвижностью — о ринхокинетизме. Дромеогнатический, или палеогнатический (у Paleognathae), тип черепа, при котором сочленение костного нёба и других костей черепа неподвижно, противопоставляют неогнатическому (у Neognathae), в который объединяют схизогнатический, десмогнатический и эгитогнатический типы.
Описание
Череп птиц принадлежит к диапсидному типу[1], он имеет два височных окна (верхнее и боковое), нижнюю височную дугу, образованную скуловой и квадратноскуловой костями, верхняя височная дуга утрачена[2]. Чтобы поместить крупный головной мозг[1], птицам потребовалась большая черепная коробка. Облегчению веса черепа способствовало приобретение клюва вместо челюсти с зубами и бесшовное соединение костей черепа у взрослых птиц[1][3]. Исследования окостенения черепа птиц обычно относятся к черепу птенцов[1].
Череп птиц часто сравнивают с черепом пресмыкающихся, у большинства из которых он тоже принадлежит к диапсидному типу, но которым не требуется такая большая черепная коробка и лёгкий клюв (вместо него у рептилий тяжёлые челюсти с зубами)[1][3]. Подвижность надклювья относительно черепа связана с усовершенствованием челюстного аппарата по сравнению с рептилиями[3].
Среди всех систем органов птиц вариации челюстного аппарата вместе с размерами и формами клюва и языка являются одними из самых разнообразных[3].
Строение


Мозговой череп
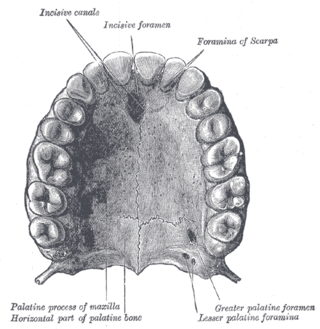
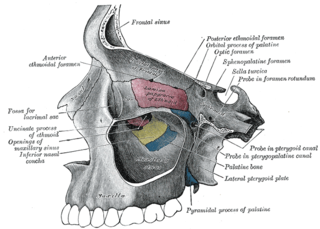
Затылочный отдел черепа состоит из верхней затылочной кости, двух боковых затылочных костей и основной затылочной кости, которые окружают затылочное отверстие. Основная затылочная кость формирует затылочный мыщелок[1], с которым связана высокая подвижность головы относительно шеи[3]. Парные теменные, лобные и носовые кости формируют крышу черепа. В слуховой капсуле развита крупная предушная кость, в передней стенке мозговой капсулы — основная клиновидная кость внизу и пара боковых клиновидных костей вверху[1]. Срединная обонятельная кость (лат. mesethmoideum) хорошо развита и формирует тонкую межглазничную перегородку. Перед глазницей расположена слёзная кость, а под ним — скуловая кость палочковидной формы, к которой сзади прирастает квадратно-скуловая кость такой же формы. Чешуйчатая кость у большинства птиц выходит на внутреннюю поверхность черепа и принимает участие в формировании его стенки. Обширный задний отдел парасфеноида — основная височная кость (лат. basitemporale) — прикрывает снизу основную клиновидную кость, а передний отдел парасфеноида — рострум (лат. rostrum parasphenoidale) — межглазничную перегородку. Сошник у птиц непарный, он прирастает к нёбным костям и расположен ниже носовых капсул[1].
Висцеральный череп
Основанием для верхней челюсти служат крыша черепа и срединная обонятельная кость[1]. Первичная верхняя челюсть состоит из трёх костей. По сравнению с рептилиями, у птиц отсутствуют поперечная кость и эпиптеригоид, рудимент которого сохранился у некоторых видов[4]. К покровной нёбной кости сзади присоединена крыловидная кость, сочленённая челюстным суставом с квадратной костью, которая может выполнять качательные движения вперёд и назад[1][4]. Она также соединена с чешуйчатой и переднеушной костями черепной коробки и может быть соединена с боковой затылочной[4]. Вторичная верхняя челюсть состоит из преднечелюстной, верхнечелюстной и носовой костей, с доминирующими размерами первой из них[1]. Костное нёбо у птиц образовано соединением нёбных отростков межчелюстных и верхнечелюстных костей, сошником, нёбными и крыловидными костями[5]. Крыловидная кость участвует в базиптеригоидном сочленении у древненёбных птиц и подвижном нёбно-крыловидном суставе у неонёбных[4].
Базиптеригоидные сочленения — скользящие сочленения с боковой поверхностью крыловидной кости, образованные выростами на боковых сторонах клиновидной кости (базиптеригоидными отростками). Они присутствуют в частности у страусоподобных птиц, тинаму, некоторых трубконосых, гусеобразных, сов. У некоторых видов, в частности пингвинов, чаек, чистиковых, они образуются на стадии эмбриона и исчезают по мере взросления птиц[5].
У основания верхней части надклювья нёбная кость и передний фрагмент крыловидной кости (гемиптеригоид) могут истончаться и образовывать соединительную перемычку (синдесмоз) или сустав[4][5]. Нёбно-крыловидный сустав позволяет крыловидной кости поворачиваться в горизонтальной плоскости, а рамке костного нёба и скуловой луги — деформироваться[6]. Сустав скользит вдоль клювовидного отростка парасфеноида[5] и позволяет широкий размах движений нёба и надклювья, чему также способствует утрата большинством птиц ограничивающего базиптеригоидного сочленения[6].
Задний край ноздри образован верхнечелюстным и межчелюстным отростками носовой кости. У птиц различают округлые и щелевидные ноздри. Округлые ноздри (голоринальный тип) образованы неглубоко разделёнными отростками, которые сходятся под широким закруглённым углом перед линией соприкосновения лобной и межчелюстной костей. Щелевидные ноздри (схизоринальный тип) образованы глубоко расщеплёнными отростками, которые сходятся под острым углом и могут заходить за линию соприкосновения лобной и межчелюстной костей[7][6]. Такое деление предложил английский зоолог Алфред Генри Гаррод[англ.] в 1873 году[6]. Округлые ноздри характерны для пингвинов, гагар, трубконосых, куриных, веслоногих, пастушковых, дроф, кукушек, ракшеобразных, дятлообразных, воробьиных, а щелевидные — для ибисов, журавлей, большинства куликов, чаек, чистиковых, голубей, рябков и некоторых других птиц[7].
Нижняя челюсть птиц (подклювье) включает задний отдел, который состоит из замещающей сочленовной кости, угловой, надугловой и предсочленовной костей, и передний отдел, состоящий из покровных костей, зубной и пластинчатой кости. По сравнению с рептилиями, у птиц отсутствует венечная кость[1]. Ветви нижней челюсти некоторых птиц могут выгибаться наружу, так как разделены на две части гибким участком. Из-за этого глотка может раскрываться значительно сильнее, что позволяет заглатывать крупную добычу[1][5]. Такое строение называется стрептогнатизмом и присутствует в частности у трубконосых, веслоногих, чаек[5].
В среднем ухе у птиц расположено стремя — звукопередающая косточка. Подвижную основу скелета стреловидного языка обеспечивают рожки, сформированные из остатков висцеральных дуг и первой жаберной дуги[1].
Кинетичность черепа

Кинетичность, или кинетизм, черепа — это способность верхней челюсти перемещаться вертикально относительно черепной коробки[1][5]. Впервые этот термин предложил датский зоолог Ян Верслюис[англ.] в 1910 году в отношении стегоцефалов и завроспид, позднее его распространили на рыб и птиц. Однако у птиц некоторые варианты кинетичности невозможны[8]. Если гибкая зона лежит у основания надклювья, то говорят о прокинетизме, а если она расположена на уровне носовых ям и верхняя челюсть обладает внутренней подвижностью — о ринхокинетизме[5][8].
В первом случае движения надклювья и квадратной кости синхронизированы, так как нижняя часть основания надклювья соединена с квадратной костью и скуловой дугой[1]. Гибкая зона образована носовыми костями на основании верхней челюсти. Между мезглазничной и носовой перегородками в этом месте находится черепнолицевая щель. Прокинетизм характерен для большинства птиц, при нём верхняя челюсть прочнее[8], ноздри как правило округлые[6].
Во втором, надклювье носит стебельчатый характер, так как основание верхней челюсти как правило прорезано щелевидной ноздрёй[1][6]. Гибкая зона захватывает предчелюстную кость и предчелюстные отростки носовых костей[8]. Ринхокинетизм является более примитивной формой[8], им обладают ржанкообразные, ибисовые и журавлеобразные[1], а также голубеобразные, тинаму и бескилевые. У вальдшнепа гибкая зона располагается в конечной трети надклювья, чтобы схватывать червей он может раскрывать только кончик клюва, полностью вонзённого в землю[8].
Кинетичность черепа помогает амортизировать рабочие усилия клюва в случае ошибок. Жёсткость верхней челюсти достигается за счёт напряжения челюстных мышц, которые вызывают кратковременные нагрузки на череп. Кроме того, кинетичностью обусловлено разнообразие производимым клювом движений, а также величина усилия на конце клюва[1].
Типы черепа
В 1867 году английский зоолог Томас Генри Гексли в работе «О классификации птиц и о таксономическом значении изменений в строении костного нёба в этом классе» (On the classification of birds, and on the taxonomic value of the modification of certain of the cranial bones observable in that Class)[9] на основе особенностей строения и взаимоотношения различных частей черепа летающих птиц определил четыре классических типа костного нёба: дромеогнатический, схизогнатический, десмогнатический, эгитогнатический[6]. Дромеогнатический, или палеогнатический (у Paleognathae), тип черепа противопоставляют неогнатическому (у Neognathae), в который иногда объединяют все другие типы[5]. Такое деление предложил английский зоолог Уильям Плейн Пайкрафт[англ.] в 1900 году, который отнёс к группе с палеогнатическим черепом бескилевых птиц. Дальнейшее определение типов вызвало оживлённую дискуссию среди учёных[6].
Дромеогнатический тип
Самый примитивный тип черепа, при котором сочленение костного нёба и других костей черепа неподвижно. У такого черепа нёбные и крыловидные кости соединены неподвижно, базиптеригоидные сочленения хорошо развиты (обычно отсутствуют у других взрослых птиц), сошник крупный, его передний конец сливается с нёбными отростками костей верхней челюсти, а задний конец расщеплён и соединяется расходящимися концами с нёбными и крыловидными костями[5][9].
Длина костного нёба в дромеогнатическом черепе неизменна, продольные перемещения сильно ограничены[10]. Неподвижное соединение не позволяет манипулировать с кормом, оно способно только для амортизации толчков[6] и простых хватательных движений[9]. Дромеогнатический тип является первым этапом эволюции кинетизма черепа у птиц[6]. У современных птиц он характерен для страусоподобных (бескилевых) птиц и тинаму[9].
Схизогнатический тип
Схизогнатический тип черепа является более древним, чем другие неонёбные, которые скорее всего возникли путём его перестройки[5]. При этом типе отсутствует соединение нёбных отросков правой и левой верхнечелюстных костей, между ними расположена длинная щель[5][6]. Сочленение нёбных и крыловидных костей подвижное, они примыкают к клювовидному отростку, отсутствует соединение сошника, заострённого спереди, с нёбными отростками верхнечелюстных костей[5].
Схизогнатический тип характерен для пингвинов, гагар, трубконосых, куриных, большинства журавлеобразных, ржанкообразных, трогонов, большинства сов, козодоевых, колибри, настоящих дятлов[5].
Десмогнатический тип
При этом типе происходит слияние нёбных отростков правой и левой верхнечелюстных костей[5][6] по средней линии, из-за чего надклювье становится более прочным и имеет трубчатую конструкцию[6]. Сошник редуцирован либо очень узкий, иногда соединяющийся задним концом с нёбными костями и клювовидным отростком, а передним — со слившимися нёбными отростками[5].
Десмогнатический тип характерен для веслоногих, голенастых, гусеобразных, дневных хищников, некоторых козодоев, ракшеобразных, якамаров, пуховок и туканов из дятлообразных[5].
Эгитогнатический тип
При этом типе нёбные отростки верхнечелюстных костей тонкие и удлинённые, они подходят под сошник, но не соприкасаются ни с ним, ни друг с другом, у широкого сошника свободный передний конец иногда раздвоён, а задний глубоко расщеплён и охватывает клювовидный отросток[5]. Функционально это схизогнатия[6].
Эгитогнатический тип характерен для трехпёрсток, стрижей, бородаток и медоуказчиков из дятлообразных, почти у всех воробьиных[5].
Морфология черепа и систематика
Гексли в 1867 году применил разделение на типы костного нёба для определения четырёх подотрядов обширной группы Carinatae[англ.]. В современной систематике этот таксономический уровень соответствует надотряду. Гексли не рассматривал родственные отношения между таксонами одной группы, но отметил низкое положение тинаму (Tinamidae)[10]. Образование костного нёба птиц рассмотрел Пайкрафт в 1900 году, который пытался выяснить пути филогенеза. Он обнаружил сходство строения мускулатуры, покровов и репродуктивной системы у птиц с дромеогнатическим нёбом — страусов, бескрылов, моа, эпиорнисов и тинаму — и объединил их в надотряд Palaeognathae, сгруппировав остальных в Neognathae. Формулировки Пайкрафта были основаны на типе соединения между palatina и pterigoidea (шовное против шаровидного), а не на подвижности сустава, что привело к многочисленным дискуссиям в этом вопросе[10].
В 1948 году МакДауэлл (McDowell) рассмотрел костное нёбо птиц как комбинацию многих мелких признаков. В его работе содержится четыре варианта нёба палеогнат и указание на наличие у неогнат сочленений, отличных от шаровидного. МакДауэлл критиковал концепцию Пайкрафта и заключил, что выделение палеогнатического и неогнатического типа необоснованно. Точка зрения МакДауэлла до сих пор поддерживается многими орнитологами[10].
Опровержение концепции Макдауэлла предоставил Хофер (Hofer) в 1955 году, развивая направление биомеханики, основанное Фершлейсом (Versluys) в 1910 году и продолженное Маринелли (Marinelli) в 1928 и 1936 годы и рядом других учёных. Они рассматривали костное нёбо как часть костно-мышечного механизма, нёбно-крыловидный сустав в котором выполняет функцию передаточного звена, от которого зависит размах, частота и сила движений верхней челюсти. Хофер подтвердил концепцию Пайкрафта в целом и значительно укрепил в частностях, но его исследования долгое время не были интерпретированы систематиками[10].
В 1970 году Юдин, наблюдая за живыми птицами в зоопарках, обнаружил различную технику захвата и заглатывания пищи у палеогнат и неогнат. Страусоподобные птицы после захвата пищи делают резкое движение шеей назад, сообщая пище ускорение вглубь пасти, а потом движение вперёд, устремляя голову навстречу заглатываемому объекту. При этом надклювье и нёбо внешне остаются неподвижными, однако небольшой сдвиг создаёт напряжённость в конструкции максиллярного сегмента. Неогнаты совершают движения головой чтобы поставить пищу в контакт с подвижным нёбом и языком, после чего они втягивают кусок вглубь пасти, совершая синхронные движения надклювья, нёба, языка, и, возможно, верхней гортани[10].
На более низких таксономических уровнях, в пределах отрядов, семейств и родов, исследования костного нёба и кинетичности черепа также дают богатый материал об эволюции птиц[10].
Существовала также точка зрения, что деление на палеогнаты и неогнаты связано с уровнем совершенствования, то есть палеогнаты сохраняют более низкий уровень развития, а неогнаты — продвинутое состояние, которое развивалось независимо. При этом ни те, ни другие, не могут считаться монофилитическими таксонами, а палеогнаты и негнаты представляют собой грады (уровни развития), а не клады[11].
Номенклатура
| Русское название | Латынь | Примечание | |
|---|---|---|---|
| Мозговой череп (neurocranium) | |||
| Затылочный отдел | |||
| Верхняя затылочная кость | supraoccipitale | ПЗП. стр. 74 | ОСЖКП. стр. 42 |
| Боковая затылочная кость | exoccipitale | ПЗП. стр. 74 | ОСЖКП. стр. 42 |
| → затылочный мыщелок | condylus occipitalis | ПЗП. стр. 74 | ОСЖКП. стр. 42 |
| → затылочное, или барабанное, крыло | ala tympanica | ПЗП. стр. 74 | ОСЖКП. стр. 42 |
| Основная затылочная кость | basioccipitale | ПЗП. стр. 74 | ОСЖКП. стр. 42 |
| Переднеушная кость | prooticum | ПЗП. стр. 75 | ОСЖКП. стр. 42 |
| → овальное окно | fenestra ovalis | ПЗП. стр. 75 | ОСЖКП. стр. 42 |
| Заднеушная кость | opisthoticum | ПЗП. стр. 75 | ОСЖКП. стр. 42 |
| → круглое окно | fenestra rotunda | ПЗП. стр. 75 | ОСЖКП. стр. 42 |
| Верхнеушная кость | epioticum | ОСЖКП. стр. 42 | |
| Клиновидный отдел | |||
| Основная клиновидная кость | basisphenoideum | ПЗП. стр. 75 | ОСЖКП. стр. 42 |
| → базиптеригоидный отросток | processus basipterygoideus | ПЗП. стр. 75 | ОСЖКП. стр. 42 |
| Боковая клиновидная кость | laterosphenoideum, также pleurosphenoideum | ПЗП. стр. 75 | ОСЖКП. стр. 42 |
| → заглазничный отросток | processus postorbitalis | ПЗП. стр. 75 | ОСЖКП. стр. 42 |
| Обонятельный отдел | |||
| Срединная обонятельная кость, или обонятельная кость | mesethomoideum | ПЗП. стр. 75 | ОСЖКП. стр. 44 |
| → межглазничная перегородка | septum interorbitale | ПЗП. стр. 75 | ОСЖКП. стр. 44 |
| → черепно-лицевая щель | fissura craniofacialis | ОСЖКП. стр. 44 | |
| Покровный череп (dermatocranium), также висцеральный череп (splanchnocranium) | |||
| Теменная кость | parietale | ПЗП. стр. 76 | ОСЖКП. стр. 44 |
| → височные гребни | linea temporalis | ОСЖКП. стр. 44 | |
| Лобная кость | frontale | ПЗП. стр. 77 | ОСЖКП. стр. 44 |
| Носовая кость | nasale | ПЗП. стр. 77 | ОСЖКП. стр. 45 |
| → лобный отросток | processus frontalis | ОСЖКП. стр. 45 | |
| → межчелюстной отросток | processus praemaxillaris | ОСЖКП. стр. 45 | |
| → челюстной отросток | processus maxillaris | ОСЖКП. стр. 45 | |
| Чешуйчатая кость | squamosum | ПЗП. стр. 77 | ОСЖКП. стр. 46 |
| → скуловой отросток | processus zygomaticus | ПЗП. стр. 77 | |
| → заднеглазничный отросток | processus orbitalis posterior, или sphenoticus | ОСЖКП. стр. 46 | |
| → височный отросток | processus temporalis | ОСЖКП. стр. 46 | |
| Предлобная кость, или слёзная кость | praefrontale | ПЗП. стр. 77 | ОСЖКП. стр. 46 |
| → надглазничный отросток | processus supraorbitalis | ОСЖКП. стр. 46 | |
| → предглазничный отросток | processus orbitalis anterior | ОСЖКП. стр. 46 | |
| Скуловая кость | jugale | ПЗП. стр. 78 | |
| Квадратно-скуловая кость | quadratojugale | ПЗП. стр. 78 | ОСЖКП. стр. 48 |
| Сошник | vomer | ПЗП. стр. 79 | ОСЖКП. стр. 44 |
| Верхняя челюсть (надклювье) | |||
| Вторичная верхняя челюсть | |||
| Предчелюстная, или межчелюстная, кость | praemaxillare | ПЗП. стр. 80 | ОСЖКП. стр. 47 |
| → лобный отросток | processus frontalis | ОСЖКП. стр. 48 | |
| → нёбный отросток | processus palatinus | ПЗП. стр. 81 | ОСЖКП. стр. 48 |
| → челюстной отросток | processus maxillaris | ОСЖКП. стр. 48 | |
| Верхнечелюстная, или челюстная, кость | maxillare | ПЗП. стр. 81 | ОСЖКП. стр. 48 |
| → нёбный отросток | processus palatinus, или maxillo-palatinum | ПЗП. стр. 81 | ОСЖКП. стр. 48 |
| → скуловой отросток | processus zygomaticus | ОСЖКП. стр. 48 | |
| Вторичная верхняя челюсть | |||
| Нёбная кость | palatinum | ПЗП. стр. 82 | ОСЖКП. стр. 44 |
| → каудолатеральная пластинка | lamina caudolateralis | ПЗП. стр. 82 | |
| Крыловидная кость | pterygoideum | ПЗП. стр. 82 | ОСЖКП. стр. 42 |
| → гемиптериоид | hemipterygoideum | ПЗП. стр. 82 | |
| Квадратная кость | quadratum | ПЗП. стр. 82 | ОСЖКП. стр. 48 |
| → ушной отросток | processus oticus | ПЗП. стр. 82 | ОСЖКП. стр. 48 |
| → глазничный отросток | processus orbitalis | ПЗП. стр. 82 | ОСЖКП. стр. 48 |
| Нижняя челюсть (подклювье) | |||
| Сочленовная кость | articulare | ПЗП. стр. 84 | ОСЖКП. стр. 53 |
| → внутренний отросток нижней челюсти | processus internus mandibulae | ПЗП. стр. 84 | ОСЖКП. стр. 53 |
| → задний отросток нижней челюсти | processus posterior mandibulae | ПЗП. стр. 84 | ОСЖКП. стр. 53 |
| Зубная кость | dentale | ПЗП. стр. 84 | ОСЖКП. стр. 53 |
| Подбородочная косточка | mento-mekkelianum | ОСЖКП. стр. 53 | |
| Пластинчатая кость | spleniale | ПЗП. стр. 84 | ОСЖКП. стр. 53 |
| Угловая кость | angulare | ПЗП. стр. 84 | ОСЖКП. стр. 53 |
| → заднеугловой отросток | processus angularis posterior | ОСЖКП. стр. 53 | |
| Надугловая кость | supraangulare | ПЗП. стр. 84 | ОСЖКП. стр. 53 |
| Предсочленовная, или покровная сочленовная, кость | praearticulare | ПЗП. стр. 84 | |
| Слуховой столбик, или стремечко | columella auris, или stapes | ПЗП. стр. 84 | |
| Подъязычный аппарат | apparatus hyobrahchialis | ПЗП. стр. 85 | ОСЖКП. стр. 54 |
Легенда:
- ПЗП — Гуртовой Н.Н., Дзержинский Ф.Я. Практическая зоотомия позвоночных. Птицы. Млекопитающие. — Москва: Высшая школа, 1992. — 414 с. — ISBN 5-06-002207-2.
- ОСЖКП — Шульпин Л.М. Орнитология (строение, жизнь и классификация птиц). — Ленинград: Издание Ленинградского государственного университета, 1940. — 555 с.
Примечания
- ↑ 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 Дзержинский, Васильев, Малахов, 2013, с. 303—306.
- ↑ Дзержинский, 2005, с. 97—98.
- ↑ 1 2 3 4 5 Коблик, 2001, с. 15—16.
- ↑ 1 2 3 4 5 Дзержинский, 2005, с. 90—92.
- ↑ 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 Карташев, 1974, Кинетизм черепа и типы строения костного нёба.
- ↑ 1 2 3 4 5 6 7 8 9 10 11 12 13 14 Дзержинский, 2005, с. 104—107.
- ↑ 1 2 Карташев, 1974, Типы скелетной основы ноздрей.
- ↑ 1 2 3 4 5 6 Дзержинский, 2005, с. 98—103.
- ↑ 1 2 3 4 Коблик, Чернова, 2017.
- ↑ 1 2 3 4 5 6 7 Юдин К. А. Классические морфологические признаки и современная систематика птиц // Русский орнитологический журнал. — 2009. — Т. 18, вып. 498. — С. 1215—1222. Архивировано 3 декабря 2019 года.
- ↑ Дзержинский, 2005, с. 21.
Литература
- Дзержинский Ф. Я., Васильев Б. Д., Малахов В. В. Зоология позвоночных. — М.: Академия, 2013. — С. 293-. — 464 с. — ISBN 978-5-7965-7971-4.
- Дзержинский Ф. Я. Сравнительная анатомия позвоночных животных. — 2-е. — М.: Аспект-Пресс, 2005. — 320 с.
- Карташев Н. Н. Систематика птиц. — М.: Высшая школа, 1974. — С. 7—. — 342 с.
- Коблик Е. А. Разнообразие птиц (по материалам экспозиции Зоологического музея МГУ). — М.: Издательство МГУ, 2001. — Т. 1. — С. 15—16. — 384 с. — ISBN 5-211-04072-4.
- Коблик Е. А., Чернова О. Древненёбные птицы. Очерки филогении, систематики, биологии, морфологии и хозяйственного использования. — Litres, 2017. — С. 7—8.
- Baumel J. J. Handbook of Avian Anatomy: Nomina Anatomica Avium. — 2nd ed. — Cambridge: Nuttall Ornithological Club, 1993. — P. 45—132.