Четвероногие
| Четвероногие | |||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 1-й ряд (земноводные): синайская агама, королевская крачка | |||||||||||||||||||||||||||||||||||||||||
| Научная классификация | |||||||||||||||||||||||||||||||||||||||||
Домен: Эукариоты Царство: Животные Подцарство: Эуметазои Без ранга: Двусторонне-симметричные Без ранга: Вторичноротые Тип: Хордовые Подтип: Позвоночные Инфратип: Челюстноротые Надкласс: Четвероногие | |||||||||||||||||||||||||||||||||||||||||
| Международное научное название | |||||||||||||||||||||||||||||||||||||||||
| Tetrapoda Hatschek & Cori, 1896 | |||||||||||||||||||||||||||||||||||||||||
| Подгруппы | |||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||
| Геохронология появился 367,5 млн лет
◄ Наше время◄ Мел-палеогеновое вымирание ◄ Триасовое вымирание◄ Массовоепермское вымирание ◄ Девонское вымирание◄ Ордовикско-силурийское вымирание◄ Кембрийский взрыв | |||||||||||||||||||||||||||||||||||||||||
Четвероно́гие, или тетрапо́ды[1] (лат. Tetrapoda), — надкласс челюстноротых из группы костных позвоночных. Характерный признак четвероногих — наличие четырёх конечностей, обычно служащих для передвижения по суше. В некоторых группах конечности существенно видоизменились для полёта (птицы, рукокрылые, птерозавры), плавания (ластоногие, китообразные, сирены, мозазавры, ихтиозавры, плезиозавры) или редуцировались (безногие земноводные, змеи и некоторые ящерицы[a]).
Определение и этимология
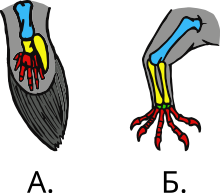
Б) Конечность ящерицы. Плечевая кость Локтевая и лучевая кости Запястье Фаланги
Процесс выделения четвероногих как группы имеет длительную и запутанную историю. В произведении Аристотеля «История животных» (книга 1; часть 5), которое датируют IV веком до н. э., автор отмечает, что некоторые животные, такие как ящерица и собака, имеют общий признак: у них четыре конечности[5].
Название Tetrapoda восходит к др.-греч. τετράπους: τετράς — «четыре», и πούς — «нога». В XIX и XX веках различные авторы применяли это или схожие с ним названия по отношению к разным таксономическим группам, чаще всего к отдельным подгруппам четвероногих в нынешнем понимании (хотя П. А. Латрей подразумевал под tétrapodes бабочек-нимфалид). Спорно, какому учёному следует приписывать авторство над таксоном Tetrapoda[5][6]. Ранее автором часто считался Э. С. Гудрич (1930), но в 2019 году Х. Дитер-Сьюз обнаружил, что наиболее раннее упоминание Tetrapoda как таксономической группы приходится на 1909 год, когда таксон выделил немецкий палеонтолог О. Йекель[5]. В 2020 году М. Лорин[англ.] пришёл к выводу, что впервые этот таксон был введён ещё раньше — в сборнике лекций по зоологии 1896 года за авторством Б. Гатчека и К. Дж. Кори[6].

С развитием филогенетической номенклатуры[англ.] встал вопрос о том, какое формальное определение должно быть закреплено за кладой Tetrapoda. Сторонники основанного на апоморфии определения полагают, что ключевым признаком четвероногих является наличие двух парных конечностей с раздельными пальцами на них (у некоторых представителей признак вторично утрачен); такой подход соответствует традиционному пониманию объёма таксона[7][8][9]. Другие авторы привязывают таксон к кроновой группе, исключая из его состава ряд ранних стволовых таксонов (таких как акантостега), что, по их мнению, обеспечит таксономическую стабильность в будущем и позволит давать более информативное описание группе, включающее признаки, которые не сохраняются в палеонтологической летописи[6][7].
По определению Ж. А. Готье и соавторов (1988), к четвероногим относится ближайший общий предок беспанцирных и амниот, а также все его потомки[10]. В томе Phylonyms (2020), с выпуском которого в силу вступил кодекс филогенетической номенклатуры «Филокодекс», даётся следующие определение клады Tetrapoda B. Hatschek & C. J. Cori 1896 [M. Laurin]: наименьшая кроновая клада, включающая человека разумного (Homo sapiens), настоящую червягу (Caecilia tentaculata), большого сирена (Siren lacertina) и суринамскую пипу (Pipa pipa); включение в определение трёх земноводных обусловлено их спорным филогенетическим положением[6][11]. Лорин (2020) предложил использовать название Stegocephali (стегоцефалы) по отношению к кладе, примерно соответствующий Tetrapoda в традиционном понимании[12][13].
Эволюционная история
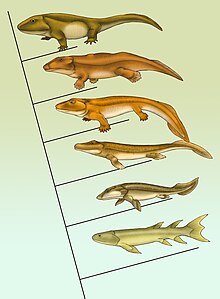
Долгое время считалось, что предками четвероногих были ископаемые кистепёрые (в традиционном понимании данной группы как одного из двух — наряду с двоякодышащими — отрядов подкласса лопастепёрых[14]; ныне последний чаще считают классом[15]). Как предполагалось, некоторые из кистепёрых, обитавшие в пересыхающих заболоченных водоёмах, выработали способность к лёгочному дыханию и научились переползать из водоёма в водоём, опираясь на парные плавники (которые постепенно трансформировались в четыре конечности с пальцами). В соответствии с этими представлениями, наиболее близкой к четвероногим группой в современном фауне являются целакантообразные, тогда как двоякодышащие связаны с наземными позвоночными в меньшей степени[16][17][18].
В конце XX — начале XXI века в эти представления были внесены корректировки. Единый отряд кистепёрых был разделён на несколько отдельных отрядов (сам термин «кистепёрые» в современной литературе используют редко[19], а если и используют, то в изменённом значении[20][21]). Согласно современным данным о филогении лопастепёрых, эта группа в начале раннего девона разошлась на две ветви. Первая — Coelacanthinimorpha — представлена отрядами Onychodontiformes[англ.] (ониходонтообразные; syn. Onychodontida, Struniiformes), и Actinistia (актинистии — целакантообразные и несколько вымерших семейств). Вторая — рипидистии (Rhipidistia) — почти сразу разделилась на две сестринские клады: Dipnomorpha[англ.] — с отрядами Porolepiformes (поролепообразные) и Dipnoi (двоякодышащие) — и Tetrapodomorpha (так что среди ныне живущих позвоночных двоякодышащие оказываются ближе к четвероногим, чем целакантообразные)[22][23][24][25][26].
Наряду с тетраподами (четвероногими), в состав тетраподоморф входят три традиционно выделяемых отряда лопастепёрых, ископаемые остатки которых известны начиная со среднего девона: Rhizodontida[англ.] (ризодонтиды), Osteolepiformes[англ.] (остеолепообразные) и Elpistostegalia[англ.] (элпистостегалии), или Panderichthyida[англ.] (пандерихтииды). Если первые два отряда включают типичных рыб, которые, однако, некоторыми особенностями строения скелета сходны с ранними четвероногими, то элпистостегалии (три рода: Panderichthys[англ.], Elpistostege и Tiktaalik) объединяют формы, переходные от рыб к четвероногим: у этих животных крокодилоподобного облика отсутствуют спинные и анальный плавники, а грудные и брюшные плавники состоят из костей, гомологичных костям конечностей четвероногих, и только членение дистальных частей плавников на пальцы ещё отсутствует[27]. Все три отряда, по современным представлениям, являются парафилетическими, из-за чего ныне их нередко разделяют на отдельные монофилетические линии. Элпистостегалии иногда сохраняются в системе как группа, в которую, помимо вымерших стволовых представителей, включают и самих четвероногих[28].
| Мясистолопастные |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| (Sarcopterygii) |


Если рассматривать неразветвлённые дистальные радиальные элементы в плавнике элпистостеги[34] (Elpistostege) как истинные пальцы, то этот таксон попадает под апоморфное определение четвероногих и, в таком случае, является древнейшим известным представителем этой группы. Клутье и коллеги (2020) восстановили этот род как самого базального четвероногого (стегоцефала), сестринского по отношению ко всем остальным апоморфным четвероногим[38]. В России древнейшие четвероногие (в широком смысле) — это Jakubsonia livnensis и Parmastega aelidae[англ.]. Якубсония обнаружена совместной группой исследователей Палеонтологического института им. А. А. Борисяка РАН и сотрудников Ливенского краеведческого музея. Возраст находки — порядка 350 млн лет назад[39]. Пармастега описана учёными из России, Швеции, Великобритании и Латвии по остаткам из отложений Сосногорской формации в республике Коми. Возраст находки — около 372 млн лет (фаменский век[англ.] поздней эпохи девонского периода)[40][41].
| ||||||||||||||||||||||||||||||
| Кладограмма по анализу Клутье и коллег (2020)[38] |
Ископаемые остатки тетрапод известны с верхнего девона. По результатам исследования А. Пирона (2011), ветви земноводных и рептилиоморф разошлись 367,5 млн лет назад, в позднем девоне[42]. Д. Марьянович (2021) пришёл к выводу, что, в зависимости от филогенетической гипотезы, кроновые четвероногие возникли примерно 335 или 350 млн лет назад[43]. В карбоновых отложениях появляются остатки завропсид и синапсид[44]. Продвинутые синапсиды — маммалиаформы ( ≈ млекопитающие в широком смысле) — появились в позднем триасе[45][46], а представители кроновой группы млекопитающих (в узком смысле) — в средней юре[47]. Возникновение наиболее специализированных завропсид — авиал ( ≈ птицы в широком смысле) — приходится на позднюю юру[48]; веерохвостые птицы (кроновая группа) появились в среднем[49] или позднем мелу[50].
Описание

По оценкам биологов (на 2011 год), надкласс четвероногих включает примерно 21 100 современных видов и ещё большее число видов вымерших (известны с начала позднего девона)[51]. Приспособившись к жизни на суше, они дышат лёгкими, воздух в которые поступает через сквозные ноздри (хоаны); отсутствие лёгких (например, у безлёгочных саламандр, дышащих через кожу) носит вторичный характер. Лёгочный тип дыхания имеют и возвратившиеся в водную среду китообразные, сирены и вымершие ихтиозавры; только приспособленные к жизни в воде личинки земноводных, а также неотенические земноводные (такие как аксолотль), дышат жабрами[52].
Подавляющее большинство четвероногих имеет две пары конечностей (их редукция или полная утрата — у безногих земноводных, змей и некоторых ящериц[a] — вторична). В исходном состоянии конечности пятипалые, иногда число пальцев уменьшается; только у позднедевонских четвероногих число пальцев ещё не вполне установилось и варьировалось от пяти до восьми (у акантостеги их было по 8, у ихтиостеги — по 7, у тулерпетона — по 6[53])[54][55]. При этом каждая конечность четвероногого представляет собой сложную систему рычагов и шарниров, подвижно сочленённых как друг с другом, так и с плечевым и тазовым поясами. Передняя конечность включает плечо (его каркас — плечевая кость), предплечье (локтевая и лучевая кости) и кисти; последняя состоит из запястья, пясти и пальцев. Аналогично, задняя конечность включает бедро (бедренная кость), голень (большая и малая берцовые кости) и стопу, а последняя по костной структуре делится на предплюсну, плюсну и фаланги пальцев[56].
Кроме того, для четвероногих характерны усложнение внутреннего скелета и зубного аппарата. В позвоночнике, помимо туловищного и хвостового, имеются также шейный и крестцовый отделы[57].
Классификация
Традиционная систематика

Слева направо: бесчелюстные, хрящевые рыбы, плакодермы (вымерли), акантоды (вымерли), костные рыбы, земноводные, пресмыкающиеся, птицы, млекопитающие.
В XVIII—XIX веках пресмыкающиеся вместе с амфибиями объединялись в группу гады — холоднокровные наземные позвоночные[59]. В эволюционной таксономии, наиболее влиятельной в XIX—XX веках, в надклассе Четвероногие традиционно выделяются 4 класса: земноводные, пресмыкающиеся, птицы и млекопитающие[60].
| Птицы (Aves) | |||||||||||||||||||||
| Земноводные (Amphibia) | Пресмыкающиеся (Reptilia) | ||||||||||||||||||||
| Млекопитающие (Mammalia) | |||||||||||||||||||||
Земноводные в данной классификации представляют собой анцестральную парафилетическую группу, а все остальные классы образуют кладу Amniota (амниоты). В свою очередь, пресмыкающиеся являются парафилетической группой по отношению к птицам и млекопитающим[61]. Актуальный на конец XX века перечень более мелких подразделений четвероногих (во многих случаях до рода включительно) можно найти в 3-м томе монографии Р. Кэрролла «Палеонтология и эволюция позвоночных» (1993)[62].
Элементы эволюционной таксономии были сохранены Красной книгой МСОП, которая признаёт все четыре класса[63].
Филогенетическая систематика

В конце XX и начале XXI веков, в свете распространения кладистики, многие специалисты отказались от приведённой выше традиционной классификации. Сторонники филогенетической систематики полагают, что любые таксоны обязательно должны быть монофилетическими, то есть включать всех потомков одного общего предка. Так, в рамках макросистемы ныне живущих организмов, предложенной в 2015 году М. Руджеро с соавторами, в составе надкласса Tetrapoda выделялись три класса: Amphibia, Reptilia и Mammalia (немаммальных синапсид авторы системы не рассматривали, а таксон Aves они включили в состав Reptilia на правах подкласса)[64][65].
Все кроновые четвероногие, в соответствии с принципами филогенетической систематики, могут быть разделены на две группы: земноводные (Amphibia; syn. Batrachomorpha) и рептилиоморфы (Reptiliomorpha)[66], или Pan-Amniota[43][67]. Современных земноводных, как правило, объединяют в кроновую группу беспанцирных (Lissamphibia)[68][69], тогда как сами земноводные оказываются тотальной группой по отношению к этому таксону. Согласно такой трактовке таксонов, земноводные являются параллельной ветвью относительно всех остальных четвероногих (рептилиоморф), а не анцестральным таксоном к ним[59][70][71][72]. При этом спорно, какие вымершие таксоны следует относить к стволовой группе земноводных. Согласно наиболее распространённой версии, беспанцирные являются ветвью темноспондильных клады диссорофоид (при включении в Temnospondyli беспанцирных первые становятся синонимом Amphibia)[72][73][74]. Некоторые авторы выводят беспанцирных из тонкопозвонковых (лепоспондилов)[75]; другие считают, что бесхвостые (Anura) и хвостатые земноводные (Urodela) произошли от темноспондилов, а безногие (Gymnophiona) — от тонкопозвонковых[76].

Ко второй ветви четвероногих — Reptiliomorpha, или Pan-Amniota, — относят кроновую группу амниот (рептилий, включая птиц, и синапсид, включая млекопитающих) и все таксоны, более близкие к амниотам, чем к земноводным. К числу рептилиоморф, не относящихся к кроновым амниотам, обычно причисляют тонкопозвонковых (Lepospondyli), хрониозухий (Chroniosuchia), эмболомеров (Embolomeri), сеймуриаморф (Seymouriamorpha) и диадектоморф (Diadectomorpha). Последние, по результатам некоторых анализов, являются настоящими амниотами — сестринской группой к синапсидам — и входят в эволюционную ветвь, в конечном итоге приведшую к млекопитающим[77][78][79]. Ещё более альтернативную точку зрения предложил М. Лорин (2010), который рассматривал эмболомеров, сеймуриаморф и темноспондильных как стволовых четвероногих, а тонкопозвонковых считал предками беспанцирных и, следовательно, представителями земноводных[75].

В ветви рептилиоморф, которая привела к амниотам, появилось характерное амниотическое яйцо с уникальным набором мембран — амнионом, хорионом и аллантоисом. Возможно, первоначально амниотическое яйцо возникло для защиты зародыша от неблагоприятных условий, а затем как экзаптация позволило амниотам размножаться вдали от воды[80].
Амниот подразделяют на две большие ветви (тотальные группы): синапсиды (Synapsida; ныне представлены млекопитающими) и завропсиды[81], или рептилии (Sauropsida / Reptilia; ныне представлены рептилиями и птицами)[82][83]. Традиционный класс пресмыкающихся (рептилий) является парафилетический группой по отношению к млекопитающим и птицам. В конце XX века среди систематиков сформировался консенсус о выделении из класса пресмыкающихся животных, которые раннее называли «зверообразными рептилиями», и объединении их вместе с млекопитающими в кладу синапсид (до этого синапсидами называли только «зверообразных»)[83].
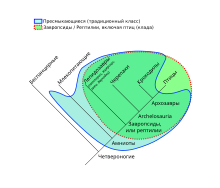
Филогенетически птицы глубоко вложены в состав рептилий, являясь при этом специализированной ветвью тероподных динозавров (Theropoda, Dinosauria)[84][85][86]. Из ныне живущих животных птицы являются ближайшими родственниками крокодилов (более близкими, чем, например, ящерицы), что стало причиной объединения этих двух таксонов в кладу архозавров (Archosauria)[87]. Таким образом, если следовать постулируемому кладистами принципу строгой монофилии таксонов, птицы могут быть признаны одной из подгрупп рептилий[64][65][83][88]. Согласно другой точке зрения, для объединения птиц и пресмыкающихся за вычетом синапсид следует использовать лишённое исторических ассоциаций название Sauropsida (завропсиды)[89][90][91]. Иногда таксон Reptilia применяют по отношению к кроновой группе (также называемой Sauria — заврии), а Sauropsida — к тотальной[20][43][92][93][94][95].
Существенную проблему систематики четвероногих составляет положение черепах. Традиционно их противопоставляли всем остальным ныне живущим рептилиям (диапсидам)[96], что хорошо согласуется с морфологическими данными[10][93][97][98][99]. Тем не менее, большинство молекулярно-генетических исследований свидетельствует в пользу того, что черепахи являются диапсидами и, вероятно, близки к архозаврам[89][100][101][102][103][104], вместе с которыми их было предложено включить в кладу Archelosauria[100][105].
Современные представления о филогенетических взаимоотношениях между основными ныне живущими группами четвероногих можно отобразить следующей кладограммой[43][66][100]:
| Четвероногие |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Биоразнообразие
Сегодня на Земле имеется большое разнообразие четвероногих, которые живут во многих средах обитания и питаются различной пищей[106]. В таблице ниже приведены сводные оценки для каждого класса четвероногих из Красной книги угрожаемых видов МСОП (2014.3), для существующих видов, которые были описаны в литературе, а также для видов, которым угрожает опасность вымирания[63].
| Глобальные сводные оценки МСОП для современных видов четвероногих на 2014 год[63] | ||||||
|---|---|---|---|---|---|---|
| Группы четвероногих | Изображение | Класс | Расчётное количество описанных видов[63] | Виды, находящиеся под угрозой исчезновения в Красной книге[63] | Виды, оцениваемые в процентах от описанных видов[63] | Оптимистичная оценка процента исчезающих видов[63] |
| Анамнии откладывают яйца в воду |  | Земноводные | 7,302 | 1,957 | 88 % | 41 % |
| Амниоты приспособлены откладывать яйца на суше | 
| Завропсиды (рептилии + включительно птицы) | 20,463 | 2,300 | 75 % | 13 % |
 | Млекопитающие | 5,513 | 1,199 | 100 % | 26 % | |
| Итого: | 33,278 | 5,456 | 80 % | ? | ||

См. также
Примечания
Комментарии
- ↑ 1 2 Безногие таксоны неоднократно возникали во многих группах ящериц (чешуйчатых). Змеи сформировались как одна из многих ветвей таких безногих ящериц и поэтому в эволюционном смысле сами являются ящерицами[2][3][4].
- ↑ Филогенетические определения:
- Eotetrapodiformes: ближайший общий предок Eusthenopteron и Ichthyostega и все его потомки[31];
- Стегоцефалы (Stegocephali): наибольшая клада, включающая Eryops megacephalus, но не Tiktaalik roseae, Panderichthys rhombolepis и Eusthenopteron foordi[12][13];
- Лабиринтодонты (Labyrinthodontia): наименьшая клада, включающая Acanthostega gunnari, Ichthyostega stensioei, Crassigyrinus scoticus, Mastodonsaurus giganteus, Baphetes kirkbyi и Anthracosaurus russelli[32][33];
- Четвероногие (Tetrapoda sensu lato): позвоночные с исходно пятипалыми конечностями[7][8][9];
- Четвероногие (Tetrapoda sensu stricto): наименьшая кроновая клада, включающая Homo sapiens, Caecilia tentaculata, Siren lacertina и Pipa pipa[6][11].
Источники
- ↑ Основы палеонтологии : Справочник для палеонтологов и геологов СССР : в 15 т. / гл. ред. Ю. А. Орлов . — М. : Наука, 1964. — Т. 12 : Земноводные, пресмыкающиеся и птицы / под ред. А. К. Рождественского, Л. П. Татаринова. — С. 23. — 724 с. — 3000 экз.
- ↑ Greene, 1997, p. 17.
- ↑ Pianka & Vitt, 2006, p. 1.
- ↑ Caldwell, 2019, chpt. 1: «Ancient Snakes, Modern Snakes: "What is a Snake?"», pp. 1—34.
- ↑ 1 2 3 Sues H.-D.[англ.]. Authorship and date of publication of the name Tetrapoda (англ.) // Journal of Vertebrate Paleontology. — 2019. — Vol. 39, iss. 1. — P. e1564758. — ISSN 0272-4634. — doi:10.1080/02724634.2019.1564758.
- ↑ 1 2 3 4 5 de Queiroz, Cantino & Gauthier, 2020, Tetrapoda B. Hatschek and C. J. Cori 1896 [M. Laurin], converted clade name, pp. 759—764.
- ↑ 1 2 3 Laurin M.[англ.], Anderson J. S. Meaning of the Name Tetrapoda in the Scientific Literature: An Exchange (англ.) // Systematic Biology : journal. — 2004. — Vol. 53, iss. 1. — P. 68—80. — ISSN 1063-5157. — doi:10.1080/10635150490264716. — PMID 14965901. — .
- ↑ 1 2 Lee M. S. Y. Stability of higher taxa in phylogenetic nomenclature — some comments on Moore (1998) (англ.) // Zoologica Scripta : journal. — 1999. — Vol. 28, iss. 3—4. — P. 361–366. — ISSN 1463-6409. — doi:10.1046/j.1463-6409.1999.00017.x. — .
- ↑ 1 2 Anderson J. S. Use of Well-Known Names in Phylogenetic Nomenclature: A Reply to Laurin (англ.) // Systematic Biology : journal. — 2002. — Vol. 51, iss. 5. — P. 822—827. — ISSN 1063-5157. — doi:10.1080/10635150290102447. — PMID 12396594. — .
- ↑ 1 2 Gauthier J., Kluge A. G., Rowe T., Benton M. J.[англ.]. The early evolution of the Amniota (англ.) // The phylogeny and classification of the tetrapods, Volume 1. Amphibians, reptiles, birds. — Oxford: Clarendon Press, 1988. — P. 103—155.
- ↑ 1 2 Tetrapoda (англ.). RegNum. Дата обращения: 21 июня 2021. Архивировано 24 июня 2021 года.
- ↑ 1 2 3 de Queiroz, Cantino & Gauthier, 2020, Stegocephali E. D. Cope 1868 [M. Laurin], converted clade name, pp. 741—745.
- ↑ 1 2 3 Stegocephali (англ.). RegNum. Дата обращения: 29 июня 2021. Архивировано 29 июня 2021 года.
- ↑ Кэрролл, т. 3, 1993, с. 190—191.
- ↑ Nelson J. S. Fishes of the World. 4th ed. — New York: John Wiley & Sons, 2006. — xx + 601 p. — ISBN 978-0-471-25031-9. — P. 459—461.
- ↑ Наумов С. П. Зоология позвоночных. — М.: Просвещение, 1982. — 464 с. — С. 81—83, 140.
- ↑ Кэрролл, т. 1, 1992, с. 189—190, 202—204.
- ↑ Медников, 1994, с. 281—283.
- ↑ Черепанов, Иванов, 2007, с. 143.
- ↑ 1 2 de Queiroz K.[англ.], Gauthier J. A. Phylogenetic Taxonomy (англ.) // Annual Review of Ecology and Systematics : journal. — 1992. — Vol. 23, iss. 1. — P. 449—480. — ISSN 0066-4162. — doi:10.1146/annurev.es.23.110192.002313. Архивировано 29 июня 2022 года.
- ↑ Benton M. J. Vertebrate Paleontology. 3rd ed. — Oxford: Blackwell Science Ltd., 2005. — 750 p. — ISBN 0-632-05637-1. — P. 393—399.
- ↑ Benton, 2015, p. 436.
- ↑ Lu J., Zhu M., Ahlberg P. E., Qiao T., Zhu Y. A Devonian predatory fish provides insights into the early evolution of modern sarcopterygians (англ.) // Science Advances : journal. — 2016. — Vol. 2, iss. 6. — P. e1600154. — ISSN 2375-2548. — doi:10.1126/sciadv.1600154. Архивировано 1 июля 2021 года. — .
- ↑ Pierce S. E., Hutchinson J. R., Clack J. A. Historical Perspectives on the Evolution of Tetrapodomorph Movement (англ.) // Integrative and Comparative Biology[англ.] : journal. — 2013. — Vol. 53, iss. 2. — P. 209—223. — ISSN 1557-7023 1540-7063, 1557-7023. — doi:10.1093/icb/ict022. — PMID 23624864. — .
- ↑ Черепанов, Иванов, 2007, с. 141—153.
- ↑ SARCOPTERYGII – coelacanths, lungfishes, tetrapods and their extinct relatives (англ.). Mikko's Phylogeny Archive. Дата обращения: 7 июля 2021. Архивировано 7 августа 2020 года.
- ↑ Черепанов, Иванов, 2007, с. 153—158.
- ↑ 1 2 Swartz B. A Marine Stem-Tetrapod from the Devonian of Western North America (англ.) // PLOS One : journal. — 2012. — Vol. 7, iss. 3. — P. e33683. — ISSN 1932-6203. — doi:10.1371/journal.pone.0033683. — PMID 22448265. Архивировано 2 апреля 2022 года. — .
- ↑ Merck J. And Now For Something Completely Different: Sarcopterygii (англ.). GEOL 431 Vertebrate Paleobiology. Дата обращения: 1 июля 2021. Архивировано 24 апреля 2021 года.
- ↑ Merck J. Elpistostegalia becomes "digitized" (англ.). GEOL 431 Vertebrate Paleobiology. Дата обращения: 1 июля 2021. Архивировано 30 июня 2021 года.
- ↑ Coates M. I., Friedman M. Litoptychus bryanti and characteristics of stem tetrapod neurocrania (англ.) // Morphology, Phylogeny and Paleobiogeography of Fossil Fishes / In Elliot D. K., Maisey J. G., Yu X., Miao D. (eds.).. — 2010. — P. 389—416. — ISBN 978-3-89937-122-2.
- ↑ 1 2 de Queiroz, Cantino & Gauthier, 2020, Labyrinthodontia R. Owen 1859 [M. Laurin], converted clade name, pp. 747—750.
- ↑ 1 2 Labyrinthodontia (англ.). RegNum. Дата обращения: 29 июня 2021. Архивировано 6 ноября 2021 года.
- ↑ 1 2 Марков и Неймарк, 2014, с. 486.
- ↑ "Павел Безносов: «Пармастега – это наш общий предок»". Республика | Издание Правительства и Государственного Совета Республики Коми. Архивировано 20 сентября 2020. Дата обращения: 1 июля 2021.
- ↑ Марков и Неймарк, 2014, с. 490.
- ↑ Любарский, 1996, с. 68.
- ↑ 1 2 Cloutier R., Clement A. M., Lee M. S. Y., Noël R., Béchard I. Elpistostege and the origin of the vertebrate hand (англ.) // Nature : journal. — 2020. — Vol. 579, iss. 7800. — P. 549—554. — ISSN 1476-4687. — doi:10.1038/s41586-020-2100-8. — PMID 32214248. — .
- ↑ Проскуряков, Евгений. В честь орловца назвали доисторическое животное // Комсомольская правда в Орле. — 2004. — № 50 (622) за 10 декабря. — С. 1.
- ↑ Beznosov P. A., Clack J. A., Lukševičs E., Ruta M., Ahlberg P. E.[англ.]. Morphology of the earliest reconstructable tetrapod Parmastega aelidae (англ.) // Nature : journal. — 2019. — Vol. 574, iss. 7779. — P. 527—531. — ISSN 1476-4687. — doi:10.1038/s41586-019-1636-y. — PMID 31645719. Архивировано 24 июня 2021 года. — .
- ↑ "У древнейших тетрапод нашли хрящевой скелет и любовь к водному образу жизни". N + 1. 2019-10-23. Архивировано 29 июня 2021. Дата обращения: 29 июня 2021.
- ↑ Pyron P. A. Divergence Time Estimation Using Fossils as Terminal Taxa and the Origins of Lissamphibia (англ.) // Systematic Biology : journal. — 2011. — Vol. 60, iss. 4. — P. 466—481. — ISSN 1063-5157. — doi:10.1093/sysbio/syr047. — PMID 21540408. — .
- ↑ 1 2 3 4 5 Marjanović D. The Making of Calibration Sausage Exemplified by Recalibrating the Transcriptomic Timetree of Jawed Vertebrates (англ.) // Frontiers in Genetics : journal. — 2021. — Vol. 12. — ISSN 1664-8021. — doi:10.3389/fgene.2021.521693. — PMID 34054911. Архивировано 10 июля 2021 года. — .
- ↑ Черепанов, Иванов, 2007, с. 162, 186, 237, 251, 264.
- ↑ Gill P. G., Purnell M. A., Crumpton N., Brown K. R., Gostling N. J. Dietary specializations and diversity in feeding ecology of the earliest stem mammals (англ.) // Nature : journal. — 2014. — Vol. 512, iss. 7514. — P. 303—305. — ISSN 1476-4687. — doi:10.1038/nature13622. — PMID 25143112. Архивировано 29 октября 2021 года. — .
- ↑ Datta P. M. Earliest mammal with transversely expanded upper molar from the Late Triassic (Carnian) Tiki Formation, South Rewa Gondwana Basin, India (англ.) // Journal of Vertebrate Paleontology. — 2005. — Vol. 25, iss. 1. — P. 200—207. — ISSN 0272-4634. — doi:10.1671/0272-4634(2005)025[0200:EMWTEU]2.0.CO;2. — .
- ↑ Bininda-Emonds O. R. P., Cardillo M., Jones K. E., MacPhee R. D. E., Beck R. M. D. The delayed rise of present-day mammals (англ.) // Nature : journal. — 2007. — Vol. 446, iss. 7135. — P. 507—512. — ISSN 1476-4687. — doi:10.1038/nature05634. — PMID 17392779. Архивировано 25 июня 2021 года. — .
- ↑ Alonso P. D., Milner A. C., Ketcham R. A., Cookson M. J., Rowe T. B. The avian nature of the brain and inner ear of Archaeopteryx (англ.) // Nature : journal. — 2004. — Vol. 430, iss. 7000. — P. 666—669. — ISSN 1476-4687. — doi:10.1038/nature02706. — PMID 15295597. Архивировано 9 февраля 2006 года. — .
- ↑ Lee M. S. Y., Cau A., Naish D.[англ.], Dyke G. J.[англ.]. Morphological Clocks in Paleontology, and a Mid-Cretaceous Origin of Crown Aves (англ.) // Systematic Biology : journal. — 2014. — Vol. 63, iss. 3. — P. 442—449. — ISSN 1063-5157. — doi:10.1093/sysbio/syt110. — PMID 24449041. Архивировано 25 февраля 2021 года. — .
- ↑ Kuhl H., Frankl-Vilches C., Bakker A., Mayr G., Nikolaus G., Boerno S. T., Klages S., Timmermann B., Gahr M. An Unbiased Molecular Approach Using 3′-UTRs Resolves the Avian Family-Level Tree of Life (англ.) // Molecular Biology and Evolution : journal. — 2021. — Vol. 38, iss. 1. — P. 108—127. — ISSN 0737-4038. — doi:10.1093/molbev/msaa191. — PMID 32781465. Архивировано 10 января 2021 года. — .
- ↑ George D., Blieck A. Rise of the earliest tetrapods: an early Devonian origin from marine environment // PLoS ONE, 2011, 6 (7): e22136. — doi:10.1371/journal.pone.0022136.. — PMID 21779385.
- ↑ Константинов, Наумов, Шаталова, 2012, с. 118, 126, 163, 327.
- ↑ Дзержинский, Васильев, Малахов, 2014, с. 236.
- ↑ Черепанов, Иванов, 2007, с. 158, 162, 164.
- ↑ Константинов, Наумов, Шаталова, 2012, с. 118.
- ↑ Михайлова, Бондаренко, 1997, с. 374—375.
- ↑ Михайлова, Бондаренко, 1997, с. 375.
- ↑ Evolutionary systematics: Spindle Diagrams (англ.). Palaeos[англ.]. Дата обращения: 5 июля 2021. Архивировано 25 февраля 2021 года.
- ↑ 1 2 de Queiroz, Cantino & Gauthier, 2020, Amphibia C. Linnaeus 1758 [M. Laurin, J. W. Arntzen, A. M. Báez, A. M. Bauer,, R. Damiani, S. E. Evans, A. Kupfer, A. Larson, D. Marjanović, H. Müller, L. Olsson, J.-C. Rage, and D. Walsh], converted clade name, pp. 765—771.
- ↑ Константинов, Наумов, Шаталова, 2012, с. 23—24.
- ↑ Черепанов, Иванов, 2007, с. 158, 162.
- ↑ Кэрролл, т. 3, 1993, с. 191—232.
- ↑ 1 2 3 4 5 6 7 The World Conservation Union. 2014. IUCN Red List of Threatened Species, 2014.3. Summary Statistics for Globally Threatened Species. Table 1: Numbers of threatened species by major groups of organisms (1996—2014) Архивная копия от 24 февраля 2015 на Wayback Machine.
- ↑ 1 2 Ruggiero M. A., Gordon D. P., Orrell T. M., Bailly N., Bourgoin T., Brusca R. C., Cavalier-Smith T., Guiry M. D., Kirk P. M. A Higher Level Classification of All Living Organisms (англ.) // PLoS ONE. — 2015. — Vol. 10, no. 4. — P. e0119248. — doi:10.1371/journal.pone.0119248. — PMID 25923521. Архивировано 14 декабря 2019 года. — .
- ↑ 1 2 Ruggiero M. A., Gordon D. P., Orrell T. M., Bailly N., Bourgoin T., Brusca R. C., Cavalier-Smith T., Guiry M. D., Kirk P. M. Correction: A Higher Level Classification of All Living Organisms (англ.) // PLoS ONE. — 2015. — Vol. 10, no. 6. — P. e0130114. — doi:10.1371/journal.pone.0130114. — PMID 26068874. Архивировано 16 июня 2021 года. — .
- ↑ 1 2 Laurin M.[англ.]. The Evolution of Body Size, Cope's Rule and the Origin of Amniotes (англ.) // Systematic Biology : journal. — 2004. — Vol. 53, iss. 4. — P. 594—622. — ISSN 1063-5157. — doi:10.1080/10635150490445706. — PMID 1360633. — .
- ↑ de Queiroz, Cantino & Gauthier, 2020, Pan-Amniota T. Rowe 2004 [M. Laurin and T. R. Smithson], converted clade name, pp. 789—792.
- ↑ de Queiroz, Cantino & Gauthier, 2020, Lissamphibia E. Haeckel 1866 [M. Laurin, J. W. Arntzen, A. M. Báez, A. M. Bauer,, R. Damiani, S. E. Evans, A. Kupfer, A. Larson, D. Marjanović, H. Müller, L. Olsson, J.-C. Rage, and D. Walsh], converted clade name, pp. 773—778.
- ↑ Lissamphibia (англ.). RegNum. Дата обращения: 7 июля 2021. Архивировано 9 июля 2021 года.
- ↑ Amphibia (англ.). RegNum. Дата обращения: 7 июля 2021. Архивировано 9 июля 2021 года.
- ↑ Gauthier J., Cannatella D. C., de Queiroz K., Kluge A. G., Rowe T. Tetrapod phylogeny (англ.) // The Hierarchy of Life / In B. Fernholm, K. Bremer, and H. Jornvall (eds.). — New York: Elsevier Science Publishers B. V., 1989. — P. 337—353.
- ↑ 1 2 Ruta M., Coates M. I. Dates, nodes and character conflict: Addressing the Lissamphibian origin problem (англ.) // Journal of Systematic Palaeontology[англ.] : journal. — 2007. — Vol. 5, iss. 1. — P. 69—122. — ISSN 1477-2019. — doi:10.1017/S1477201906002008. — .
- ↑ Schoch R. R. The putative lissamphibian stem-group: phylogeny and evolution of the dissorophoid temnospondyls (англ.) // Journal of Paleontology[англ.] : journal. — 2019. — Vol. 93, iss. 1. — P. 137—156. — ISSN 1937-2337 0022-3360, 1937-2337. — doi:10.1017/jpa.2018.67. — .
- ↑ Coates M. I., Ruta M., Friedman M. Ever Since Owen: Changing Perspectives on the Early Evolution of Tetrapods (англ.) // Annual Review of Ecology, Evolution, and Systematics : journal. — 2008. — Vol. 39, iss. 1. — P. 571—592. — ISSN 1543-592X. — doi:10.1146/annurev.ecolsys.38.091206.095546. Архивировано 9 июля 2021 года. — .
- ↑ 1 2 Laurin, 2010, p. 133.
- ↑ Fröbisch N. B., Schoch R. R. Testing the Impact of Miniaturization on Phylogeny: Paleozoic Dissorophoid Amphibians (англ.) // Systematic Biology. — 2009. — Vol. 58, iss. 3. — P. 312—327. — ISSN 1063-5157. — doi:10.1093/sysbio/syp029. — .
- ↑ Berman D. S. Diadectomorphs, amniotes or not? (англ.) // New Mexico Museum of Natural History and Science Bulletin. — 2013. — Vol. 60. — P. 22—35.
- ↑ Klembara J., Hain M., Ruta M., Berman D. S., Pierce S. E. Inner ear morphology of diadectomorphs and seymouriamorphs (Tetrapoda) uncovered by high-resolution x-ray microcomputed tomography, and the origin of the amniote crown group (англ.) // Palaeontology[англ.] : journal. — 2020. — Vol. 63, iss. 1. — P. 131—154. — ISSN 1475-4983. — doi:10.1111/pala.12448. — .
- ↑ Ruta M., Clack J. A., Smithson T. R. A review of the stem amniote Eldeceeon rolfei from the Viséan of East Kirkton, Scotland (англ.) // Earth and Environmental Science Transactions of The Royal Society of Edinburgh : journal. — 2020. — Vol. 111, iss. 3. — P. 173—192. — ISSN 1755-6929 1755-6910, 1755-6929. — doi:10.1017/S1755691020000079. — .
- ↑ Starck J. M., Stewart J. R., Blackburn D. G. Phylogeny and evolutionary history of the amniote egg (англ.) // Journal of Morphology : journal. — 2021. — Vol. 282, iss. 7. — P. 1080—1122. — ISSN 1097-4687. — doi:10.1002/jmor.21380. — PMID 33991358. Архивировано 12 июля 2021 года. — .
- ↑ Ford D. P., Benson R. B. J. A redescription of Orovenator mayorum (Sauropsida, Diapsida) using high-resolution μCT, and the consequences for early amniote phylogeny (англ.) // Papers in Palaeontology : journal. — 2019. — Vol. 5, iss. 2. — P. 197—239. — ISSN 2056-2802. — doi:10.1002/spp2.1236. Архивировано 9 июля 2021 года. — .
- ↑ Sues, 2019, p. 20—21.
- ↑ 1 2 3 Modesto S. P., Anderson J. S. The Phylogenetic Definition of Reptilia (англ.) // Systematic Biology. — 2004. — Vol. 53, iss. 5. — P. 815—821. — ISSN 1063-5157. — doi:10.1080/10635150490503026. — PMID 15545258. Архивировано 22 февраля 2021 года. — .
- ↑ Gauthier J. A. Saurischian monophyly and the origin of birds (англ.) // The Origin of Birds and the Evolution of Flight. Memoirs of the California Academy of Sciences / In K. Padian[англ.] (ed.). — 1986. — Vol. 8. — P. 1—55. Архивировано 16 августа 2019 года. — .
- ↑ Gauthier J. A., de Queiroz K.[англ.]. Feathered dinosaurs, flying dinosaurs, crown dinosaurs, and the name Aves (англ.) // New perspectives on the origin and early evolution of birds: proceedings of the International Symposium in Honor of John H. Ostrom. — 2001. — P. 7—41. Архивировано 2 июня 2020 года. — .
- ↑ Weishampel, Dodson & Osmólska, 2004, chpt. 11: "Basal Avialae" by K. Padian[англ.], pp. 210—231.
- ↑ Nesbitt S. J.[англ.]. The Early Evolution of Archosaurs: Relationships and the Origin of Major Clades (англ.) // Bulletin of the American Museum of Natural History[англ.] : journal. — 2011. — Vol. 352. — P. 1—292. — ISSN 1937-3546 0003-0090, 1937-3546. — doi:10.1206/352.1. Архивировано 1 июля 2019 года. — .
- ↑ Villela O. A. F. Reptilia vs. Sauropsida (исп.) // Revista Lationoamericana de Herpetología. — 2021. — P. 239—245. — ISSN 2594-2158. Архивировано 29 июня 2021 года.
- ↑ 1 2 Tzika A. C., Helaers R., Schramm G., Milinkovitch M. C. Reptilian-transcriptome v1.0, a glimpse in the brain transcriptome of five divergent Sauropsida lineages and the phylogenetic position of turtles (англ.) // EvoDevo. — 2011. — Vol. 2, iss. 1. — P. 19. — ISSN 2041-9139. — doi:10.1186/2041-9139-2-19. — PMID 21943375. — .
- ↑ Organ C. L., Janes D. E. Evolution of sex chromosomes in Sauropsida (англ.) // Integrative and Comparative Biology[англ.] : journal. — 2008. — Vol. 48, iss. 4. — P. 512—519. — ISSN 1540-7063. — doi:10.1093/icb/icn041. — PMID 21669812. — .
- ↑ Merck J. The Reptilian Stem (англ.). GEOL 431 Vertebrate Paleobiology. Дата обращения: 30 июня 2021. Архивировано 25 апреля 2021 года.
- ↑ Gauthier J. A.., Prothero D. R., Schoch R. M. The diversification of the amniotes (англ.) // Major features of vertebrate evolution. — Knoxville: Paleontological Society[англ.], 1994. — P. 129—159.
- ↑ 1 2 Laurin M., Reisz R. R. A reevaluation of early amniote phylogeny (англ.) // Zoological Journal of the Linnean Society[англ.] : journal. — 1995. — Vol. 113, iss. 2. — P. 165—223. — ISSN 0024-4082. — doi:10.1111/j.1096-3642.1995.tb00932.x. Архивировано 2 июня 2021 года. — .
- ↑ de Queiroz, Cantino & Gauthier, 2020, Reptilia C. Linnaeus 1758 [M. Laurin and R. R. Reisz], converted clade name, pp. 1027—1031.
- ↑ Reptilia (англ.). RegNum. Дата обращения: 7 июля 2021. Архивировано 9 мая 2021 года.
- ↑ Кэрролл, т. 1, 1992, гл. 10: "Примитивные амниоты и черепахи" (пер. Лебедев О. А.), с. 235—264.
- ↑ Reisz R. R. The origin and early evolutionary history of amniotes (англ.) // Trends in Ecology & Evolution : journal. — 1997. — Vol. 12, iss. 6. — P. 218—222. — ISSN 0169-5347. — doi:10.1016/s0169-5347(97)01060-4. — PMID 21238045. — .
- ↑ Lyson T. R., Bever G. S., Bhullar B.-A. S., Joyce W. G., Gauthier J. A. Transitional fossils and the origin of turtles (англ.) // Biology Letters : journal. — 2010. — Vol. 6, iss. 6. — P. 830—833. — ISSN 1744-9561. — doi:10.1098/rsbl.2010.0371. — PMID 20534602. — .
- ↑ Lichtig A. J., Lucas S. G. Chinlechelys from the Upper Triassic of New Mexico, USA, and the origin of turtles (англ.) // Palaeontologia Electronica[англ.] : journal. — 2021. — Vol. 24, iss. 1. — P. 1—49. — ISSN 1935-3952 1094-8074, 1935-3952. — doi:10.26879/886. Архивировано 9 июля 2021 года. — .
- ↑ 1 2 3 Crawford N. G., Parham J. F., Sellas A. B., Faircloth B. C., Glenn T. C., Papenfuss T. J., Henderson J. B., Hansen M. H., Simison W. B. A phylogenomic analysis of turtles (англ.) // Molecular Phylogenetics and Evolution : journal. — 2015. — Vol. 83. — P. 250—257. — ISSN 1055-7903. — doi:10.1016/j.ympev.2014.10.021. — PMID 25450099. — .
- ↑ Shen X.-X., Liang D., Wen J.-Z., Zhang P. Multiple Genome Alignments Facilitate Development of NPCL Markers: A Case Study of Tetrapod Phylogeny Focusing on the Position of Turtles (англ.) // Molecular Biology and Evolution[англ.]. — 2011. — Vol. 28, iss. 12. — P. 3237—3252. — ISSN 1537-1719 0737-4038, 1537-1719. — doi:10.1093/molbev/msr148. — PMID 21680872. — .
- ↑ Chiari Y., Cahais V., Galtier N., Delsuc F. Phylogenomic analyses support the position of turtles as the sister group of birds and crocodiles (Archosauria) (англ.) // BMC Biology[англ.] : journal. — 2012. — Vol. 10, iss. 1. — P. 65. — ISSN 1741-7007. — doi:10.1186/1741-7007-10-65. — PMID 22839781. — .
- ↑ Crawford N. G., Faircloth B. C., McCormack J. E., Brumfield R. T., Winker K. More than 1000 ultraconserved elements provide evidence that turtles are the sister group of archosaurs (англ.) // Biology Letters : journal. — 2012. — Vol. 8, iss. 5. — P. 783—786. — ISSN 1744-957X. — doi:10.1098/rsbl.2012.0331. — PMID 9933444. Архивировано 11 марта 2021 года. — .
- ↑ Wang Z., Pascual-Anaya J., Zadissa A., Li W., Niimura Y. The draft genomes of soft–shell turtle and green sea turtle yield insights into the development and evolution of the turtle–specific body plan (англ.) // Nature Genetics : journal. — 2013. — Vol. 45, iss. 6. — P. 701—706. — ISSN 1061-4036. — doi:10.1038/ng.2615. — PMID 23624526. Архивировано 25 мая 2021 года. — .
- ↑ Joyce W. G., Anquetin J. Cadena E.-A., Claude J., Danilov I. G. A nomenclature for fossil and living turtles using phylogenetically defined clade names (англ.) // Swiss Journal of Palaeontology : journal. — 2021. — Vol. 140, iss. 1. — P. 5. — ISSN 1664-2384. — doi:10.1186/s13358-020-00211-x. Архивировано 3 апреля 2021 года. — .
- ↑ Sahney S., Benton M. J. and Ferry P. A. Links between global taxonomic diversity, ecological diversity and the expansion of vertebrates on land (англ.) // Biology Letters : journal. — Royal Society Publishing, 2010. — August (vol. 6, no. 4). — P. 544—547. — doi:10.1098/rsbl.2009.1024. — PMID 20106856. — PMC 2936204. Архивировано 6 ноября 2015 года.
Литература
На русском языке
- Кэрролл Р. Палеонтология и эволюция позвоночных: В 3-х т. Т. 1. — М.: Мир, 1992. — 280 с. — ISBN 5-03-001819-0.
- Любарский Г. Ю. Архетип, стиль и ранг в биологической систематике. — М.: Т-во научн. изданий КМК, 1996. — 432 с.
- Марков А., Неймарк Е. Эволюция. Классические идеи в свете новых открытий. — М.: ACT: Korpus, 2014. — 656 с. — ISBN 978-5-17-083218-7.
- Медников Б. М. Биология: формы и уровни жизни. — М.: Просвещение, 1994. — 415 с. — ISBN 5-09-004384-1.
- Дзержинский Ф. Я., Васильев Б. Д., Малахов В. В. Зоология позвоночных. 2-е изд. — М.: Академия, 2014. — 464 с. — ISBN 978-5-4468-0459-7.
- Кэрролл Р. Палеонтология и эволюция позвоночных: В 3 т. Т. 3. — М.: Мир, 1993. — 312 с. — ISBN 5-03-001819-0.
- Шмальгаузен И. И. Происхождение наземных позвоночных. — М.: Наука, 1964. — 273 с.
- Константинов В. М., Наумов С. П., Шаталова С. П. Зоология позвоночных. 7-е изд. — М.: Издат. центр «Академия», 2012. — 448 с. — ISBN 978-5-7695-9293-5.
- Михайлова И. А., Бондаренко О. Б. Палеонтология. Ч. 1. — М.: Изд-во Моск. ун-та, 1997. — 448 с. — ISBN 5-211-03841-X.
- Черепанов Г. О., Иванов А. О. Палеозоология позвоночных. — М.: Издат. центр «Академия», 2007. — 352 с. — ISBN 978-5-7695-3104-0.
На английском языке
- Benton M. J.[англ.] Vertebrate palaeontology[англ.] (англ.). — 4th ed. — Wiley-Blackwell, 2015. — 480 p. — ISBN 978-1-118-40755-4. — ISBN 978-1-118-40684-7.
- The Dinosauria[англ.] (англ.) / D. B. Weishampel, P. Dodson, H. Osmólska , eds. — 2nd ed. — Berkeley: University of California Press, 2004. — 880 p. — ISBN 978-0-520-25408-4.
- Phylonyms: A Companion to the PhyloCode (англ.) / de Queiroz K.[англ.], Cantino P. D., Gauthier J. A., eds. — Boca Raton: Taylor & Francis Group, CRC Press, 2020. — 1352 p. — ISBN 978-1-138-33293-5.
- Caldwell M. W. The Origin of Snakes: Morphology and the Fossil Record (англ.). — Boca Raton, FL: CRC Press, 2019. — 326 p. — ISBN 978-1-4822-5134-0.
- Vaughan T. A., Ryan J. M., Czaplewski N. J. Mammalogy. 5th ed. — Sudbury, Massachusetts: Jones & Bartlett Learning, 2011. — 750 p. — ISBN 978-0-7636-6299-5.
- Laurin M.[англ.] How Vertebrates Left the Water (англ.). — University of California Press, 2010. — ISBN 978-0-520-26647-6.
- Sues H.-D.[англ.] The Rise of Reptiles: 320 Million Years of Evolution (англ.). — Baltimore: Johns Hopkins University Press, 2019. — 385 p. — ISBN 9781421428680. — ISBN 1421428687.
- Greene H. Snakes: The Evolution of Mystery in Nature. (англ.). — Berkeley, CA: University of California Press, 1997. — 351 p. — ISBN 978-0-520-20014-2.
- Pianka E. R., Vitt L. J. Lizards: Windows to the Evolution of Diversity (англ.). — Berkeley, CA: University of California Press, 2006. — 333 p. — ISBN 978-0-520-24847-2.
- Clack J. A. Gaining ground: the origin and evolution of tetrapods. — Bloomington, Indiana, USA: Indiana University Press, 2012. — xvi + 523 с. — ISBN 978-0-253-35675-8.
| Таксономия | |
|---|---|
| В библиографических каталогах |
| Elginerpetontidae |
| 
| |||||
|---|---|---|---|---|---|---|---|
| Acanthostegidae | |||||||
| Ichthyostegidae | |||||||
| Tulerpetidae | |||||||
| Аистоподы |
| ||||||
| Ватчеериды |
| ||||||
| Colosteidae |
| ||||||
| Crassigyrinidae | |||||||
| Baphetoidea |
| ||||||
| Прочие |
| ||||||
| Четвероногие | |||||||