
Дезоксирибонуклеи́новая кислота́ (ДНК) — макромолекула, обеспечивающая хранение, передачу из поколения в поколение и реализацию генетической программы развития и функционирования организмов. Молекула ДНК хранит биологическую информацию в виде генетического кода, состоящего из последовательности нуклеотидов. ДНК содержит информацию о структуре различных видов РНК и белков.

Транскри́пция — происходящий во всех живых клетках процесс синтеза РНК с использованием ДНК в качестве матрицы; перенос генетической информации с ДНК на РНК.

Рекомбинация — перераспределение генетического материала путём разрыва и соединения разных молекул, приводящее к появлению новых комбинаций генов или других нуклеотидных последовательностей. В широком смысле слова включает в себя не только рекомбинацию между молекулами ДНК, но и перекомбинацию (сортировку) генетического материала на уровне целых хромосом или ядер, а также обмен плазмидами между клетками.

Репарация — особая функция клеток, заключающаяся в способности исправлять химические повреждения и разрывы в молекулах ДНК, повреждённых при нормальном биосинтезе ДНК в клетке или в результате воздействия физических или химических реагентов. Осуществляется специальными ферментными системами клетки. Ряд наследственных болезней связан с нарушениями систем репарации.
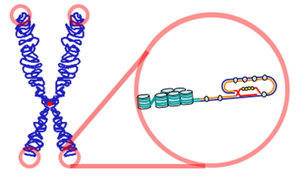
Теломе́ры — концевые участки хромосом. Теломерные участки хромосом характеризуются отсутствием способности к соединению с другими хромосомами или их фрагментами и выполняют защитную функцию.
Теломераза — фермент, добавляющий особые повторяющиеся последовательности нуклеотидов ДНК к 3'-концу цепи ДНК на участках теломер, которые располагаются на концах хромосом в эукариотических клетках. Теломеры содержат уплотнённую ДНК и стабилизируют хромосомы. При каждом делении клетки теломерные участки укорачиваются. Существование механизма, компенсирующего укорочение теломер (теломеразы), было предсказано в 1973 году А. М. Оловниковым.

Ku — белок, связывающийся с двуцепочечными разрывами в структуре ДНК. Ku необходим для репарации ДНК по пути негомологичного соединения концов (NHEJ). Эволюционно Ku консервативен от бактерий до человека. Древний бактериальный Ku является гомодимером, эукариотический гомолог Ku — гетеродимер, состоящий из двух полипептидов — Ku70 и Ku80, молекулярная масса этих полипептидов составляет 70 и 80 кДа, соответственно. Субъединицы Ku образуют корзиноподобную структуру, закрепляющуюся на конце молекулы ДНК. После связывания Ku может скользить по цепочке ДНК, на конец которой могут нанизываться новые молекулы Ku. У высших эукариот Ku образует комплекс с каталитической субъединицей ДНК-зависимой протеинкиназы (DNA-PKcs) и образует полную ДНК-зависимую протеинкиназу, DNA-PK. Ku, по-видимому, функционирует как молекулярный остов, к которому прикрепляются другие белки, принимающие участие в процессе NHEJ.
ATM — серин/треониновая протеинкиназа, которая рекрутируется и активируется двунитевыми разрывами ДНК. Эта киназа фосфорилирует несколько ключевых белков, которые инициируют остановку клеточного цикла, запускают репарацию ДНК или апоптоз. Некоторые из этих белков, в том числе p53, Сhk2 и вариантный гистон H2AX являются опухолевыми супрессорами. Белок кодируется геном ATM.

Нуклеоли́н — белок, имеющийся у высших эукариот. У человека он кодируется геном NCL, расположенным на 2-й хромосоме в локусе 2q37.1.
Ко́мплекс MRN — белковый комплекс, состоящий из белков MRE11, Rad50 и Nbs1. У эукариот комплекс MRN/Х играет важную роль в первоначальном процессинге двуцепочечных разрывов в ДНК перед репарацией по пути гомологичной рекомбинации или негомологичного соединения концов. Комплекс MRN эффективно связывает двунитевые разрывы как in vitro, так и in vivo и может служить для фиксации концов разрыва перед репарацией. Комплекс MRN также участвует в активации киназы контрольной точки ATM в ответ на повреждение ДНК. В активации ATM комплексом MRN задействовано образование коротких одноцепочечных олигонуклеотидов эндонуклеазой Mre11.

SMC5 или белок структурной поддержки хромосом номер пять — это белок, который у человека кодируется геном SMC5.
Белок Est3 — компонент теломеразного комплекса. Играет важную роль в её деятельности.

Ku70 — белок, в человеческом организме кодируемый геном XRCC6.

Негомологи́чное соедине́ние концо́в, или негомологи́чное воссоедине́ние концо́в — один из путей репарации двунитевых разрывов в ДНК. Негомологичным этот процесс называется потому, что повреждённые концы цепи соединяются лигазой напрямую, не нуждаясь в гомологичном шаблоне, в отличие от процесса гомологичной рекомбинации. NHEJ существенно менее точен, чем гомологичная рекомбинация, и часто он приводит к потере нуклеотидов, транслокациям или слиянию теломер, при этом последние два могут являться признаками опухолевой клетки. NHEJ обнаружен у представителей всех царств живой природы, кроме того, в клетках млекопитающих он служит основным путём репарации двуцепочечных разрывов.

Гомологи́чная рекомбина́ция, или о́бщая рекомбина́ция, — тип генетической рекомбинации, во время которой происходит обмен нуклеотидными последовательностями между двумя похожими или идентичными хромосомами. Это наиболее широко используемый клетками способ устранения двух- или однонитевых повреждений ДНК. Гомологичная рекомбинация также создаёт разнообразие комбинаций генов во время мейоза, обеспечивающих высокий уровень наследственной изменчивости, что, в свою очередь, позволяет популяции лучше адаптироваться в ходе эволюции. Различные штаммы и виды бактерий и вирусов используют гомологичную рекомбинацию в процессе горизонтального переноса генов.

RecBCD — фермент бактерии Escherichia coli, инициирующий процесс гомологичной рекомбинации при репарации двух- и одноцепочечных повреждений молекулы ДНК, возникающих в результате ионизирующего излучения, ошибок в процессе репликации, ошибок в работе эндонуклеаз или в результате окислительного стресса. RecBCD — это одновременно и хеликаза, раскручивающая двойную спираль ДНК, и нуклеаза, которая её разрезает.

АДФ-рибозили́рование — химическая реакция присоединения одного или нескольких остатков АДФ-рибозы к белку. Это обратимая посттрансляционная модификация, которая играет важную роль во многих клеточных процессах, таких как передача сигнала, репарация ДНК, регуляция экспрессии генов и апоптоз. Неправильное АДФ-рибозилирование наблюдается при некоторых формах рака. Многие бактериальные токсины, такие как холерный токсин и дифтерийный токсин, влияют на АДФ-рибозилирование.
Повреждение клеток представляет собой различные изменения стресса, которым клетка подвергается из-за внешних, а также внутренних изменений окружающей среды. Среди других причин это может быть связано с физическими, химическими, инфекционными, биологическими, алиментарными или иммунологическими факторами. Повреждение клеток может быть обратимым или необратимым. В зависимости от степени повреждения клеточный ответ может быть адаптивным, и, по возможности, гомеостаз восстанавливается. Гибель клеток происходит, когда тяжесть повреждения превышает способность клетки к самовосстановлению. Гибель клеток зависит как от продолжительности воздействия вредного стимула, так и от тяжести причинённого ущерба. Гибель клеток может происходить путём некроза или апоптоза.

Контрольные точки клеточного цикла — это механизмы контроля в эукариотическом клеточном цикле, которые обеспечивают его правильное развитие. Каждая контрольная точка служит потенциальной точкой завершения клеточного цикла, во время которой оцениваются условия клетки, при этом продвижение через различные фазы клеточного цикла происходит только при соблюдении благоприятных условий. В клеточном цикле есть много контрольных точек, но три основных из них: контрольная точка G1, также известная как контрольная точка начала или ограничения или основная контрольная точка; контрольная точка G2/M; и переход от метафазы к анафазе, также известный как контрольная точка веретена. Прохождение через эти контрольные точки в значительной степени определяется активацией циклин-зависимых киназ регуляторными белковыми субъединицами, называемыми циклинами, различные формы которых продуцируются на каждой стадии клеточного цикла для контроля специфических событий, происходящих в нём.

Контрольная точка мейотической рекомбинации отслеживает мейотическую рекомбинацию во время мейоза и блокирует вход в метафазу I, если рекомбинация не обрабатывается эффективно.
