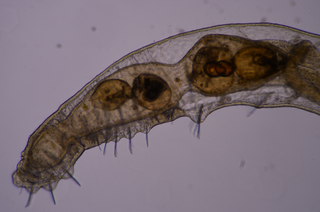
Щетинки спиральных

Щетинки, или хеты (лат. chaetae, от греч. χαίτη «волос») — хитиновые выросты кутикулы, характерные для кольчатых червей[1][2] и плеченогих[3][4]. Каждая щетинка продуцируется единственной клеткой, называемой хетобластом, который представляет собой впячивание кожного эпидермиса[1]. Для многих кольчатых червей строение щетинок является одним из наиболее значимых таксономических признаков; анатомически щетинки представляют собой одну из наиболее изученных структур полихет[2].
Терминология
Зачастую в специализированной литературе под щетинками подразумеваются исключительно структуры особого строения, присущие кольчатым червям и плеченогим[3][5]. Однако, этим же словом в зоологии могут обозначаться и другие тонкие выросты тела, в частности, у членистоногих. При этом, никакой гомологии между этими образованиями нет, а их устройство принципиально отличается: такие «щетинки» (также называемые шпорами) представляют собою крупные волоски, то есть колпачковидные выросты экзокутикулы трихогенных клеток[6].
Совокупость щетинок называется щетинковым аппаратом. Существуют различные методики описания конфигурации щетинкового аппарата. Так, для представителей люмбрицид (земляных червей) принято обозначение щетинок буквами латинского алфавита (так как один сегмент несёт по 4 щетинки с каждой стороны, они обозначаются как щетинки a, b, c, d от наиболее брюшной к наиболее спинной). Промежутки между щетинками обозначаются двумя буквами (например, bc — промежуток между щетинками b и c, dd — между щетинками d справа и слева). Методики И. И. Малевича и Т. С. Перель предполагают оценку соотношений длин промежутков aa:ab:bc:cd:dd посредством окуляр-микрометра (так, для Lumbricus terrestris указывается соотношение 4:1:3,5:0,75:16). Иванцив В. В. предлагает использовать для подобных измерений сегменты 4—6 как имеющие наиболее округлую форму посредством курвиметра либо топографического транспортира (вместо окуляр-микрометра) для повышения точности измерения[7].
Строение щетинки
Принципиальное строение щетинок у всех групп кольчатых червей и плеченогих одинаково[8]. Щетинка представляет собою хитиновое образование, продуцируемое специальной клеткой эпителия, называемой хетобластом и погружённой в слой эпителия посредством инвагинации (впячивания). Хетобласт на апикальном полюсе покрыт большим числом микроворсинок, выделяющих наружу микрофибриллярные трубочки[9] β-хитина (CMT), сшиваемые в единое целое молекулами склеропротеина — белка, выделяемого клетками, выстилающими инвагинацию[1]. Помимо этого в составе щетинки содержатся неорганические вещества, придающие ей дополнительную твёрдость[2].
Каждая щетинка состоит из большого числа продольных каналов, причём центральные каналы обычно шире периферических и имеют большие просветы, заполненные люменом. Структура вещества щетинок варьирует от мелковолокнистой до зернистой, снаружи бывает покрыта очень тонким электронноплотным слоем. Жёсткость щетинок очень варьирует, достигая особо высоких значений в зубцах крючковидных щетинок[2].
Бо́льшая часть щетинок выдаётся за пределы тела и контактирует с внешней средой. Каждая щетинка выходит из собственного фолликула. Нередко эти фолликулы снабжены большим количеством разнообразных желёз, усиливающих выделение слизистого секрета[10]. Несколько таких фолликулов, принадлежавших щетинкам единого пучка или ряда, формируют щетинковый мешок, подстилаемый общим базальным матриксом. Щетинки соединяются с базальным матриксом с помощью полудесмосом. Каждая группа щетинок (а иногда и отдельные наиболее специализированные щетинки) обладает собственной мускулатурой, прикрепляющейся к матриксу[2]. Среди мышц, обслуживающих щетинки, различают ретракторы и протракторы, а иногда и другие типы мышц, например, для Stylaria показано также наличие передних и задних сгибателей и вращателей[10].
Часто внутри щетинкового мешка помимо уже развитых щетинок можно встретить добавочные щетинки более мелкого размера, вырастающие на замену основным щетинкам, являющимся в целом недолговечным образованием[10].
Разнообразие щетинок
Форма и количество щетинок в пределах пучка крайне вариабельны и являются важными таксономическими признаками. Для одних только олигохет показано не менее 10 типов щетинок, среди которых волосные, веерные, равнозубчатые, игловидные, палочковидные, крючковидные и половые. Исходными формами щетинок считаются волосные и двузубчатые. Для первых характерно отсутствие зубцов и плавное сужение к дистальному концу. Среди волосных щетинок выделяют гладкие и оперённые, то есть покрытые волосками меньшего размера. Оперённость может быть как одно-, так и двусторонней. Двузубчатые щетинки имеют два зубца, дистальный и проксимальный, часто изогнутые по отношению к главной оси щетинки; иногда между ними наблюдается более мелкий промежуточный зубец. У некоторых видов один из зубцов (реже оба) претерпевает редукцию; если оба зубца редуцированы и притом дистальный конец щетинки расширен, её называют весловидной. Щетинки могут иметь как прямую, так и изогнутую форму вплоть до S-образной. Иногда в срединной части щетинки присутствует расширение, получившее название узелка.
У олигохет различают половые щетинки, которые в дистальной части могут быть полыми либо бороздчатыми. Среди половых щетинок выделяют сперматекальные (располагаются вблизи семяприёмников) и пениальные (располагаются среди мужских половых отверстий[10]).
У эррантных полихет выделяют особый тип щетинок, называемых ацикулами. Ацикулы имеют крупный размер и не выступают за пределы тела червя, располагаясь внутри параподий, выступая в роли их скелетных и поддерживающих элементов. Подобные структуры, по-видимому, независимо появляются у представителей семейства Psammodrilidae[2].
Щетинки нотоподии и невроподии полихет могут заметно различаться[5]. Аналогичным образом, часто друг относительно друга специализируются и щетинки брюшных и спинных пучков олигохет[10].
Распространение
Щетинки характерны для большинства видов кольчатых червей. Они встречаются у многих полихет, как седентарных (сидячих), так и эррантных (блуждающих), выполняя и локомоторную, и питательную функцию, участвуя в фильтрации[5]. Наличие щетинок достоверно показано и для эхиурид несмотря на утрату последними сегментации. Эхиуриды обладают 1 парой брюшных щетинок (гомологичных щетинкам невроподий), для некоторых групп также характерно наличие двух дополнительных полукруглых пучков щетинок на заднем конце тела[11]. У олигохет параподии отсутствуют, но щетинки сохраняются практически во всех группах (исключение представляет род Achaeta, сохраняющий только щетинковые железы при редукции самих щетинок, а также некоторые другие малощетинковые: Parergodrillus, Rheomorpha, ряд Lumbriculidae)[10]. Для пиявок характерна полная редукция щетинок: они сохраняются лишь у древних (щетинконосных) пиявок, и то лишь на 5 наиболее передних сегментах тела[12].

Помимо кольчатых червей, щетинки присутствуют у плеченогих, у которых они располагаются по краю мантии[13]. Они лишены внешнего эмалеподобного слоя и притом имеют зазубрины и шипики, однако в целом устроены почти идентично щетинкам кольчатых червей[8]. Щетинки кольчатых червей и плеченогих считаются гомологичными образованиями[8]. В пользу этого говорит экспрессия одного и того же гомеобоксного гена из группы Hox Post1 в щетинконосных мешках этих беспозвоночных. Предполагается, что форониды и мшанки, более родственные плеченогим по сравнению с кольчатыми червями, вторично утратили щетинки вместе с геном Post1[3].
Гипотетически предполагается также наличие щетинок у ископаемых брюхоногих моллюсков. Такие моллюски, описанные в качестве вида Pelagiella exigua, были обнаружены в Пенсильвании и относятся к окаменелостям раннего кембрия[13]. Их щетинки имеют длину до 1,3 мм и сгруппированы в 2 пучка по 50 штук в каждом. Насчёт происхождения этих щетинок имеется две точки зрения. Согласно первой из них, их наличие отражает предковое состояние, характерное для спиральных животных (Lophotrochozoa) в целом. Альтернативное мнение предполагает, что щетинки Pelagiella возникли заново, но являются глубокими гомологами таковых у кольчатых червей и брахиопод, образовавшись за счёт того же набора генов[9]. Проблемой обеих версий является неясный таксономический статус рода Pelagiella, по-видимому, сборного. Есть основания предполагать, что на самом деле многие виды, отнесённые к нему, включая Pelagiella exigua, представляют собой остатки полихет из группы Sabellida, на что указывают детали структуры их минерализованных трубок (или раковин)[14].
Так или иначе, такая ключевая преадаптация для возникновения щетинок, как возможность синтеза хитиновых микрофибриллярных трубочек (CMT), по-видимому, характерна для большинства спиральных животных и утрачивается лишь у форонид, сипункулид и отдельных групп моллюсков[13]. Образования, в той или иной степени напоминающие щетинки, сохраняются у хитонов и личинок осьминогов, а также наблюдаются у вымерших виваксий[9].
См. также
Примечания
- ↑ 1 2 3 Kristian Fauchald. Worms, Annelida (англ.) // Encyclopedia of Biodiversity. — Elsevier, 2013. — P. 408–419. — ISBN 978-0-12-384720-1. — doi:10.1016/b978-0-12-384719-5.00170-2. Архивировано 2 апреля 2019 года.
- ↑ 1 2 3 4 5 6 Harald Hausen. Chaetae and chaetogenesis in polychaetes (Annelida) (англ.) // Morphology, Molecules, Evolution and Phylogeny in Polychaeta and Related Taxa / T. Bartolomaeus, G. Purschke. — Berlin/Heidelberg: Springer-Verlag, 2005. — Vol. 179. — P. 37–52. — ISBN 978-1-4020-2951-6. — doi:10.1007/1-4020-3240-4_4.
- ↑ 1 2 3 Ludwik Gąsiorowski. Phoronida—A small clade with a big role in understanding the evolution of lophophorates (англ.) // Evolution & Development. — 2023-04-29. — ISSN 1520-541X. — doi:10.1111/ede.12437. Архивировано 12 августа 2023 года.
- ↑ Scott Santagata. Brachiopoda (англ.) // Evolutionary Developmental Biology of Invertebrates 2 / Andreas Wanninger. — Vienna: Springer Vienna, 2015. — P. 263–277. — ISBN 978-3-7091-1870-2. — doi:10.1007/978-3-7091-1871-9_12.
- ↑ 1 2 3 Christopher J. Glasby, Patricia A. Hutchings, Kristian Fauchald, Gregory W. Rouse & Robin S. Wilson. Polychaetes & Allies (англ.) / Editors: P.L.Beesley, G.J.B.Ross, C.J.Glasby. — Australian Biological Resources Study/CSIRO Publishing, 2000. — Vol. 4A. — 465 p. — (Fauna of Australia). — ISBN Hardcover ISBN 0 643 06571 7. — ISBN Hardcover ISBN 978 0 643 06571 0. Архивировано 12 августа 2023 года.
- ↑ Шванвич Б. Н. Курс общей энтомологии. Введение в изучение строения и функций тела насекомых. — М.—Л.: Советская наука, 1949. — С. 362. — 899 с. Архивировано 12 августа 2023 года.
- ↑ Иванцив В. В. Щетинковый аппарат дождевых червей (Oligochaeta: Lumbricidae) и его использование в диагностике таксонов видовой группы // Вестник зоологии : Журнал. — Киев: Наукова думка, 1989. — Июнь (№ 3). — С. 76—77. — ISSN 0884-5604.
- ↑ 1 2 3 Carsten Luter, Thomas Bartolomaeus. The phylogenetic position of Brachiopoda-a comparison of morphological and molecular data (англ.) // Zoologica Scripta. — 1997-07. — Vol. 26, iss. 3. — P. 245–253. — ISSN 0300-3256. — doi:10.1111/j.1463-6409.1997.tb00414.x. Архивировано 12 августа 2023 года.
- ↑ 1 2 3 Раннекембрийские предки улиток уже обзавелись раковиной, но еще обладали органами кольчатых червей • Новости науки. «Элементы». Дата обращения: 12 августа 2023. Архивировано 12 августа 2023 года.
- ↑ 1 2 3 4 5 6 Чекановская О. В. Водные малощетинковые черви фауны СССР / под ред. Е. Н. Павловского. — М.—Л.: Издательство АН СССР, 1962. — 411 с. Архивировано 18 июля 2023 года.
- ↑ Ekin Tilic, Janina Lehrke, Thomas Bartolomaeus. Homology and Evolution of the Chaetae in Echiura (Annelida) // PLOS ONE. — 2015-03-03. — Т. 10, вып. 3. — С. e0120002. — ISSN 1932-6203. — doi:10.1371/journal.pone.0120002.
- ↑ Лукин Е. И. Пиявки пресных и солоноватых водоёмов. Фауна СССР. — Л.: Наука, 1976. — Т. 1. — С. 195—207. — 484 с. Архивировано 10 марта 2023 года.
- ↑ 1 2 3 Roger D. K. Thomas, Bruce Runnegar, Kerry Matt. Pelagiella exigua , an early Cambrian stem gastropod with chaetae: lophotrochozoan heritage and conchiferan novelty (англ.) // Palaeontology / Julia Sigwart. — 2020-07. — Vol. 63, iss. 4. — P. 601–627. — ISSN 0031-0239. — doi:10.1111/pala.12476. Архивировано 12 августа 2023 года.
- ↑ Ed Landing, Gerd Geyer, Igor A. Jirkov, Stefano Schiaparelli. Lophotrochozoa in the Cambrian evolutionary radiation and the Pelagiella problem (англ.) // Papers in Palaeontology / Julia Sigwart. — 2021-11. — Vol. 7, iss. 4. — P. 2227–2244. — ISSN 2056-2799. — doi:10.1002/spp2.1396. Архивировано 14 августа 2023 года.