Щетинконосные пиявки
| Щетинконосные пиявки | |
|---|---|
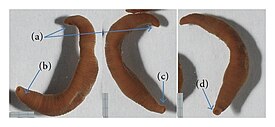 Внешний вид Acanthobdella peledina из реки Чечуй: (a) передний конец тела, (b) задняя присоска; (c) удлинённый задний конец тела; (d) задняя присоска в стандартном состоянии | |
| Научная классификация | |
Домен: Эукариоты Царство: Животные Подцарство: Эуметазои Без ранга: Двусторонне-симметричные Без ранга: Первичноротые Без ранга: Спиральные Тип: Кольчатые черви Класс: Поясковые черви Подкласс: Пиявки Инфракласс: Древние пиявки (Archihirudinea) Отряд: Acanthobdellida Подотряд: Acanthobdelliformes Семейство: Щетинконосные пиявки | |
| Международное научное название | |
| Acanthobdellidae Livanow, 1905 | |
| Роды | |
| |
Щетинконосные пиявки[1], или щетинковые пиявки[2], или древние пиявки[3] (лат. Acanthobdellidae) — семейство пиявок (Hirudinea), относящееся к монотипному отряду Acanthobdellida[4]. Являются сестринской группой по отношению к настоящим пиявкам. Семейство включает 2 вида: Acanthobdella peledina и Paracanthobdella livanowi[5].
Название
Название семейства образовано от названия типового представителя — Acanthobdella, в свою очередь происходящего от др.-греч. άκανθος ‘шип, щетинка’ и βδέλλᾰ ‘пиявка’, отражающего наличие щетинок (вооружения) на переднем конце тела[2].
Подотряд Acanthobdelliformes (Cios, de Carle, Świątek, Tessler & Utevsky, 2023) был выделен в целях большего номенклатурного соответствия другим группам пиявок сходного ранга (Oceanobdelliformes, Glossiphoniiformes и др.), образованных посредством латинского суффикса -formes ‘подобные’[5].
Название Archihirudinea («древние пиявки», от др.-греч. ἀρχι- ‘начальный’ и лат. hirudinea ‘пиявки’) было введено Е. И. Лукиным (1956) как обозначение подкласса для противопоставления настоящим пиявкам (Euhirudinea)[3].
В английском языке применительно к щетинконосным пиявкам иногда используются словосочетания «leech-like annelids» ‘кольчатые черви, похожие на пиявок’[6] и «leech-like salmonid parasites» ‘паразиты лососёвых рыб, похожие на пиявок’[7], подчёркивающие их систематическую обособленность и плезиоморфность, не позволяющую считать их пиявками в полном смысле этого слова.
Описание
Внешнее строение
Пиявки небольшого размера (длина тела 1,0—14,9 мм, ширина 0,4—4,4 мм). Масса тела варьирует от 0,2 до 42 мг[8], после насыщения — до 234 мг[3]. Paracanthobdella livanowi существенно мельче, чем Acanthobdella peledina. Тело типичной для пиявок формы, неправильно веретенобразное, сужается к концам, наибольшей ширины достигает около третьей четверти длины, позади срединных сегментов. Передний конец тела утончён сильнее заднего, не несёт настоящей присоски[3]. У Paracanthobdella livanowi с возрастом передний конец развивается, образуя углубление, по строению аналогичное присоске других пиявок, но развитое слабее[6]. Задняя присоска образована 4 сегментами, развита сравнительно слабо, ориентирована перпендикулярно продольной оси тела[8].
Взрослые пиявки имеют тёмную окраску, причём передние два кольца каждого сегмента имеют бурый цвет, а два задних — жёлтый. Молодые особи окрашены светлее, имеют монотонно-желтоватую пигментацию[8]. Фиксированные экземпляры теряют пигментацию, приобретая беловатую или желтоватую окраску[3].
Тело сегментированное, разделено на 29 (Acanthobdella peledina) или 30 (Paracanthobdella livanowi) сегментов[8] (по другим данным, число сегментов у обоих видов равняется 31; различие объясняется сложностью в разделении сегментов, слагающих заднюю присоску)[5]. Полный сегмент состоит из 4 колец. При этом сегменты I и II состоят из 1 кольца, сегмент III — из двух, а сегменты XXII—XXV со спинной стороны сохраняют 4 кольца, но на брюшной состоят только из трёх (борозда неполная)[Комментарий 1].

На 5 передних сегментах тела располагаются щетинки крючковидной формы, функционально замещающие переднюю присоску, способствуя прикреплению пиявки к телу хозяина. На каждом сегменте располагается 8 щетинок, сгруппированных в 4 пучка по две, причём первый ряд щетинок окружает ротовое отверстие. Каждая щетинка имеет в центральной части утолщение, напоминающее узелок на щетинках олигохет. Представители Paracanthobdella livanowi демонстрируют бо́льшую степень специализации щетинок по сравнению с Acanthobdella peledina[9]. Щетинки имеют коричневатую окраску, темнеющую ближе к дистальному концу[5]. Помимо функционирующих щетинок при внимательном рассмотрении в пучках можно отметить щетинки меньшего размера, которые являются замещающими в случае повреждения основных[3].
Имеется 3 пары глаз, расположенные на III, IV и V сегментах соответственно. По сравнению с глазами настоящих пиявок, развиты слабо; при фиксации разрушаются и потому могут быть трудно обнаружимы[3]. Эти глаза занимают на теле маргинальное или даже боковое положение, что является признаком, свойственным в первую очередь хищным пиявкам, активно ищущим свою добычу[8].
Гистология
Кожные покровы представлены однослойным эпителием толщиною 15—20 мкм и состоят из основных, чувствительных и железистых клеток. Некоторые клетки эпителия погружены в подлежащие ткани. Кутикула типичная для кольчатых червей, состоит из базальной кутикулы и эпикутикулы.
Мускулатура состоит из одного слоя кольцевых, 2 слоёв диагональных и нескольких слоёв продольных мышечных волокон (число последних варьирует). Волокна обладают явной косой исчерченностью и погружены во внеклеточный матрикс, переходящий в паренхиматозные соединительные ткани. Каждое волокно содержит единственное ядро размером 15×20×80 мкм, расположенное в саркоплазме и занимающее центральное положение. Т-система отсутствует.
Целом хорошо развит (что отличает древних пиявок от настоящих, для которых свойственна редукция целома). В каждом сегменте располагается отдельная целомическая полость. Её размер меньше такового у олигохет в связи с развитием мускулатуры. Целомические полости передних 5 сегментов сливаются в области вокруг глотки, так что между ними остаются лишь боковые септы. Целомические полости остальных сегментов более автономны, однако септы всё равно остаются неполными в областях дорзальнее и вентральнее кишечника. Мезентерии развиты слабо. В сегментах задней присоски целом отсутствует[10].
Анатомия
Пищеварительная система сквозная, состоит из ротового отверстия, короткого пищевода, желудка и кишечника, а также анального отверстия. Для Acanthobdella peledina показано менее ясное разграничение отделов пищеварительной системы по сравнению с Paracanthobdella livanowi, имеющей чёткую границу между пищеводом (сравнительно более длинным) и желудком, а также разделением желудка на 6 отделов (7 отдел формирует кишечник, 8 отдел — прямую кишку). Прямая кишка соединяется с анальными железами не вполне ясного назначения. Пищеварительная система хорошо развита уже у ювенильных особей и со взрослением практически не претерпевает изменений[8]. Наиболее активное пищеварение происходит в кишечнике[11].
Нервная система включает в себя мозг (надглоточную ганглионарную массу), подглоточную ганглионарную массу, цепочку свободных ганглиев и анальную ганглионарную массу в области задней присоски. Брюшная нервная цепочка включает 21 ганглий и начинается от последнего сегмента, несущего щетинки. Строение ганглия типично для представителей пиявок: он состоит из 6 групп нейронов, среди которых 2 непарных (передний и задний медиовентральные) и 4 парных (переднелатеральные и заднелатеральные). Каждый ганглий соединяется с периферией тела посредством образования 3 пар боковых корешков, причём первый иннервирует 1 кольцо в сегменте, второй — 2 кольцо, а третий разветвляется, иннервируя 3 и 4 кольца. Подглоточная ганглионарная масса состоит из 24 групп нейронов, что указывает на сращение 4 ганглиев; аналогично, надглоточная ганглионарная масса, будучи состоящей из 12 групп нейронов, соответствует 2 ганглиям[10].
Гермафродиты. Имеют 2 раздельных половых отверстия (гонопоры), причём мужская гонопора расположена между IX и X сегментами, а женская — между 3 и 4 кольцами X сегмента (расстояние между гонопорами составляет 3 кольца)[3]. Обе гонопоры расположены вентрально на медиальной линии, разделяющей правую и левую половину тела; открываются на пояске (мужская гонопора — между II и III сегментами пояска, женская — на III сегменте). На границе пояска и уросомы располагается семяприёмник[5]. Мужская половая система включает семенные мешки, выносящие каналы, семяпроводы, семенные пузырьки и семяизвергательные каналы, расширяющиеся в прямые рога атриума (конечной части мужских половых путей, представляющей собой расширение в виде камеры[3]). Женская половая система включает яйцевые мешки (их длина вдвое меньше длины семенных мешков у Acanthobdella peledina и приблизительно равна длине семенных мешков у Paracanthobdella livanowi), яйцеводы и вагину[8]. Семенные мешки заполнены сперматоцитами, сперматидами и сперматозоидами на разных стадиях развития[10].
Жизненный цикл
Копуляция происходит при низких температурах (экспериментальные данные показали, что повышение температуры с 4 °С до 10 °С понижает частоту копуляции). Коконы образуются спустя неделю после обмена половыми клетками. В образовании коконов участвует секрет, производимый железами пояска; во время откладки кокона пиявка прикрепляется к твёрдому субстрату. Коконы достигают длины 5 мм и содержат от 13 до 33 яиц. Вылупление происходит спустя 7 месяцев[9]. Новорождённые пиявки вскоре прикрепляются к телу хозяина (чаще всего, лососёвой рыбы) и вплоть до сезона размножения находятся на нём. Половое созревание происходит осенью, свойственна протандрия (как, впрочем, и для настоящих пиявок). В октябре половая система достигает наибольшего развития, что сопровождается значительным увеличением массы тела. Жизненный цикл одногодичный[3].
Образ жизни и экология
В отличие от экологически сходных рыбьих пиявок, не способны плавать, в связи с чем передвигаются исключительно путём ползания. Выделяют также позы покоя и настороженности, идентичные таковым у рыбьих пиявок. Потревоженные пиявки способны изменять форму переднего конца тела, образуя шейное сужение таким образом, что передние сегменты несколько вздуваются и отделяются от остального тела; щетинки при этом принимают положение, при котором их крючки перпендикулярны поверхности тела.
Исключительно пресноводные формы. Являются оксифилами (обитают в водоёмах с высоким содержанием кислорода). Особые дыхательные движения, свойственные многим другим пиявкам, не показаны[3].

Эктопаразиты рыб, обитающих в реках, родниках и олиготрофных озёрах. Наиболее частыми хозяевами становятся лососёвые рыбы: кумжа (Salmo trutta), кунджа (Salvelinus leucomaenis), палия (Salvelinus lepechini), белый голец (Salvinellus albus), голец Шмидта (Salvelinus schmidti), дальнеозёрский голец (Salvelinus krogiusae), мальма (Salvelinus malma), нельма (Stenodus leucichthys nelma), тугун (Coregonus tugun), пелядь (Coregonus peled), чир (Coregonus nasus), сибирский сиг, или пыжьян (Coregonus pidschian), муксун (Coregonus muksun), омуль (Coregonus autamnalis), сибирская ряпушка (Coregonus sardinella), валёк (Prosopium cylindraceum), острорылый ленок (Brachymystax lenok), кижуч (Oncorhynchus kisutch) и нерка (Oncorhynchus nerka). Помимо этого, щетинконосные пиявки встречаются на представителе близкого семейства хариусовых рыб — сибирском хариусе (Thymallus arcticus), а также, изредка, на налимах (Lota lota) и трёхиглых колюшках (Gasterosteus aculeatus)[3][6][12].
В некоторых регионах щетинконосные пиявки являются массовыми паразитами: так, в нижнем течении Енисея Acanthobdella peledina поражает 9,3 % популяции нельмы, 7,7 % — мускуна, 4,6 % — чира, причём на одной особи нельмы могли одновременно питаться до 26 пиявок (интенсивность заражения кумжи доходила до 36 пиявок на особь)[3].
Чаще всего щетинконосных пиявок находят в основании плавников рыб, в особенности спинного, однако также они могут встречаться по всей поверхности тела. Поражение щетинконосными пиявками может наносить рыбам значительный вред: после питании пиявки на их теле остаются кровоточащие язвочки, а при сильном заражении нельмы происходит практически полное разрушение плавников, а также повреждение мышц[3]. После смерти рыбы в течение часа пиявки могут оставаться на её поверхности и питаться не только кровью, но и разрушающимися тканями[9]. Способность питаться тканями рыб помимо крови связывают со специфическим набором пищеварительных ферментов, свойственных щетинконосным пиявкам: показано, что по составу и активности ферментов они наименее похожи на остальных пиявок, включая рачьих. Так, для Acanthobdellidae не показана активность валин-ариламидазы, цистин-ариламидазы, трипсина и химотрипсина, а также некоторых липаз и гликозидаз, свойственных настоящим пиявкам[11].
Вероятно, в отсутствие рыб щетинконосные пиявки могут переходить к хищному образу жизни и питаться донными беспозвоночными: так, в желудке 2 представителей вида Paracanthobdella livanowi были обнаружены остатки личинок комаров-звонцов (Chironomidae) и стрекоз (Odonata)[8].
Географическое распространение
Обитают в Голарктике. Acanthobdella peledina обнаружена на севере Скандинавского полуострова, в Финляндии, на севере России от Онежского озера до Чукотки, а также на Аляске. Среди наиболее южных местообитаний — водоёмы Северо-Байкальского и Станового нагорий[3][8]. Ареал Paracanthobdella livanowi существенно у́же и включает лишь Чукотку, Камчатку и несколько местообитаний в Тауйской губе (Магаданская область)[6].
Несмотря на широкое распространение, ареал щетинконосных пиявок представляется разорванным: так, они отсутствуют в бассейне Северной Двины и немногочисленны в бассейне Оби. Предположительно, нынешние местообитания являются осколками сплошного плейстоценного ареала, наиболее обширного во время ледникового периода, когда содержание кислорода в водоёмах и водотоках было максимальным для четвертичного периода[3].
Примечательно, что щетинконосные пиявки практически не встречаются в одних водоёмах с рыбьими пиявками, по-видимому, не выдерживая конкуренции. При этом на Чукотке обнаружены водоёмы, в которых были встречены одновременно оба вида щетинконосных пиявок[3].
Таксономия
Систематическое положение
Древние пиявки являются одними из наиболее базальных представителей группы Hirudinea (пиявок в широком смысле). Более базальной группой являются лишь рачьи пиявки (Branchiobdellida), длительное время считавшиеся таксоном, лишь конвергентно напоминающим пиявок, а на самом деле занимающим особое положение среди поясковых кольчатых червей (на настоящий момент показано, что рачьи пиявки являются группой, сестринской ко всем остальным пиявкам)[5][10].
| Пиявки (Hirudinea) |
| ||||||||||||
Классификация
Семейство Acanthobdellidae включает всего 2 вида:
- Acanthobdella peledina (Grube, 1851)
- Paracanthobdella livanowi (Epstein, 1966)
Вопрос о том, какова таксономическая дистанция между данными видами, поднимался неоднократно. Изначально они были описаны В. М. Эпштейном в составе одного рода Acanthobdella, однако затем он же выделил Paracanthobdella livanowi вначале в отдельный род, а затем и в отдельное семейство Paracanthobdellidae на основании морфологических данных, прежде всего наличия передней присоски. Современные молекулярные исследования говорят о различии этих видов на уровне родов, а не семейств, в связи с чем выделение Paracanthobdellidae не поддерживается[5].
Генетика
На настоящий момент не было проведено полногеномного секвенирования щетинконосных пиявок. Известен полный митохондриальный геном: он включает 36 генов (13 из них кодируют белки; 2 — рибосомальную РНК,12S и 16S; 21 — транспортные РНК; 1 ген является предполагаемым контрольным регионом) и имеет длину 14 640 пар оснований[13].
Примечания
Комментарии
- ↑ Применительно к пиявкам принято обозначать номера сегментов римскими цифрами, номера колец в составе сегментов — арабскими.
Ссылки
- ↑ Измайлова Н. Л., Ляшенко О. А., Антонов И. В. Биотестирование и биоиндикация состояния водных объектов: учебно-методическое пособие к лабораторным работам по прохождению учебной (ознакомительной) практики. — СПб.: СПбГТУРП, 2014. — 52 с. Архивировано 26 февраля 2020 года.
- ↑ 1 2 Лукин, 1987, с. 512—513.
- ↑ 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 Лукин Е. И. Пиявки пресных и солоноватых водоёмов. Фауна СССР. Пиявки. — Л.: Наука, 1976. — Т. 1. — С. 11. — 484 с. Архивировано 10 марта 2023 года.
- ↑ Acanthobdellidae (англ.). www.gbif.org. Дата обращения: 29 июля 2023. Архивировано 29 июля 2023 года.
- ↑ 1 2 3 4 5 6 7 Danielle Božena de Carle, Łukasz Gajda, Aleksander Bielecki, Stanisław Cios, Joanna M Cichocka, Heidi E Golden, Andrew D Gryska, Sergey Sokolov, Marina Borisowna Shedko, Rune Knudsen, Serge Utevsky, Piotr Świątek, Michael Tessler. Recent evolution of ancient Arctic leech relatives: systematics of Acanthobdellida // Zoological Journal of the Linnean Society. — 2022-07-16. — Т. 196, вып. 1. — С. 149–168. — ISSN 0024-4082. — doi:10.1093/zoolinnean/zlac006.
- ↑ 1 2 3 4 Serge Y. Utevsky, Sergei G. Sokolov, Marina B. Shedko. New records of the chaetiferous leech-like annelid Paracanthobdella livanowi (Epshtein, 1966) (Annelida: Clitellata: Acanthobdellida) from Kamchatka, Russia (англ.) // Systematic Parasitology. — 2013-01. — Vol. 84, iss. 1. — P. 71–79. — ISSN 0165-5752. — doi:10.1007/s11230-012-9390-7.
- ↑ Michael Tessler, Danielle de Carle, Madeleine L. Voiklis, Olivia A. Gresham, Johannes S. Neumann, Stanisław Cios, Mark E. Siddall. Worms that suck: Phylogenetic analysis of Hirudinea solidifies the position of Acanthobdellida and necessitates the dissolution of Rhynchobdellida (англ.) // Molecular Phylogenetics and Evolution. — 2018-10. — Vol. 127. — P. 129–134. — doi:10.1016/j.ympev.2018.05.001. Архивировано 16 июня 2022 года.
- ↑ 1 2 3 4 5 6 7 8 9 Aleksander Bielecki, Joanna Maria Cichocka, Iwona Jeleń, Piotr Świątek, Bartosz Jan Płachno, Dorota Pikuła. New data about the functional morphology of the chaetiferous leech-like annelids Acanthobdella peledina (Grube, 1851) and Paracanthobdella livanowi (Epshtein, 1966) (Clitellata, Acanthobdellida): MORPHOLOGY OF A. peledina AND P. livanowi (англ.) // Journal of Morphology. — 2013-12. — P. n/a–n/a. — doi:10.1002/jmor.20235. Архивировано 29 июля 2023 года.
- ↑ 1 2 3 U. Kutschera & V. M. Epshtein. Nikolaj A. Livanow (1876 – 1974) and the living relict Acanthobdella peledina (Annelida, Clitellata) (англ.) // Annals of the History and Philosophy of Biology : журнал. — 2006. — Vol. 11. — P. 85—98. Архивировано 29 июля 2023 года.
- ↑ 1 2 3 4 G. Purschke, W. Westheide, D. Rohde, R. O. Brinkhurst. Morphological reinvestigation and phylogenetic relationship of Acanthobdella peledina (Annelida, Clitellata) (англ.) // Zoomorphology. — 1993-06. — Vol. 113, iss. 2. — P. 91–101. — ISSN 0720-213X. — doi:10.1007/BF00403087.
- ↑ 1 2 J. M. Cichocka, A. Bielecki, P. Świątek, I. Jabłońska-Barna, J. Kobak, J. Hildebrand, M. Dmitryjuk, W. Strużyński, M. Rost-Roszkowska. The activity of hydrolytic enzymes in the digestive system of Acanthobdellida, Branchiobdellida and Hirudinida (Annelida, Clitellata) – considerations on similarity and phylogeny // The European Zoological Journal. — 2021-01-01. — Т. 88, вып. 1. — С. 26–43. — ISSN 2475-0263. — doi:10.1080/24750263.2020.1851402.
- ↑ Irina A. Kaygorodova, Elena V. Dzyuba, Nikolay M. Pronin. Leech-Like Parasites (Clitellata, Acanthobdellida) Infecting Native and Endemic Eastern Siberian Salmon Fishes (англ.) // The Scientific World Journal. — 2012-05-02. — Vol. 2012. — P. e652827. — ISSN 2356-6140. — doi:10.1100/2012/652827. Архивировано 29 июля 2023 года.
- ↑ Alexander Bolbat, Gennady Vasiliev, Irina Kaygorodova. The first mitochondrial genome of the relic Acanthobdella peledina (Annelida, Acanthobdellida) (англ.) // Mitochondrial DNA Part B. — 2020-07-02. — Vol. 5, iss. 3. — P. 3282–3283. — ISSN 2380-2359. — doi:10.1080/23802359.2020.1814173. Архивировано 29 июля 2023 года.
Литература
- Лукин Е. И. Пиявки пресных и солоноватоводных водоёмов. — Фауна СССР.. — Л.: Наука, 1976. — С. 195—207.
- Лукин Е. И. Класс Пиявки (Hirudinea) // Жизнь животных. В 7 т. / гл. ред. В. Е. Соколов. — 2‑е изд., перераб. — М. : Просвещение, 1987. — Т. 1 : Простейшие. Кишечнополостные. Черви / под ред. Ю. И. Полянского. — С. 512—513. — 576 с. : ил.















