Дыхательная цепь переноса электронов, также электрон-транспортная цепь (сокр. ЭТЦ, англ. ETC, Electron transport chain) — система трансмембранных белков и переносчиков электронов, необходимых для поддержания энергетического баланса. ЭТЦ поддерживает баланс за счёт переноса электронов и протонов из НАД∙Н и ФАДН2 в акцептор электронов. В случае аэробного дыхания акцептором может быть молекулярный кислород (О2). В случае анаэробного дыхания акцептором могут быть NO3−, NO2−, Fe3+, фумарат, диметилсульфоксид, сера, SO42−, CO2 и т. д. ЭТЦ у прокариот локализована в ЦПМ, у эукариот — на внутренней мембране митохондрий. Переносчики электронов расположены в порядке уменьшения сродства к электрону, то есть по своему окислительно-восстановительному потенциалу, где у акцептора самое сильное сродство к электрону. Поэтому транспорт электрона на всём протяжении цепи протекает самопроизвольно с выделением энергии. Выделение энергии в межмембранное пространство при переносе электронов происходит ступенчато, в виде протона (H+). Протоны из межмембранного пространства попадают в протонную помпу, где наводят протонный потенциал. Протонный потенциал преобразуется АТФ-синтазой в энергию химических связей АТФ. Сопряжённая работа ЭТЦ и АТФ-синтазы носит название окислительного фосфорилирования.

Окисли́тельное фосфорили́рование — метаболический путь, при котором энергия, образовавшаяся при окислении питательных веществ, запасается в митохондриях клеток в виде АТФ. Хотя различные формы жизни на Земле используют разные питательные вещества, АТФ является универсальным соединением, в котором запасается энергия, необходимая для других метаболических процессов. Почти все аэробные организмы осуществляют окислительное фосфорилирование. Вероятно, широкому распространению этого метаболического пути способствовала его высокая энергетическая эффективность по сравнению с анаэробным брожением.
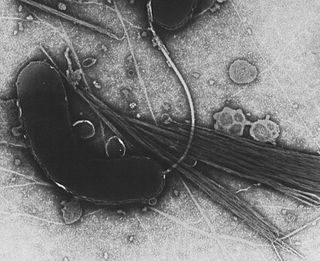
Холерный вибрион — вид грамотрицательных факультативно анаэробных подвижных бактерий рода вибрионов. Открыт Филиппо Пачини в 1854 году и названа им Filippo Pacini bacillum. Вибрион был повторно и независимо обнаружен Робертом Кохом в 1883 году. Vibrio cholerae серогрупп О1 и О139 являются возбудителями холеры и отнесены ко II группе патогенности.
Протеинкина́зы — подкласс ферментов киназ (фосфотрансфераз). Протеинкиназы модифицируют другие белки путём фосфорилирования остатков аминокислот, имеющих гидроксильные группы или гетероциклической аминогруппы гистидина.
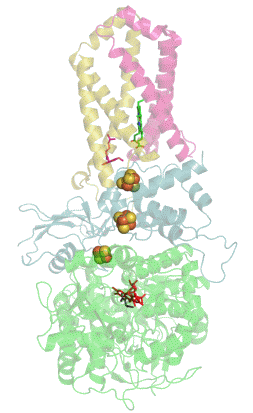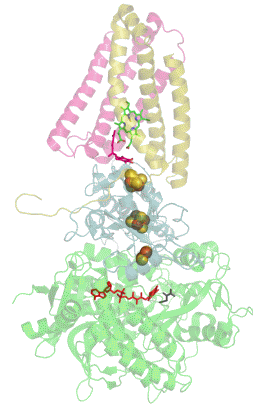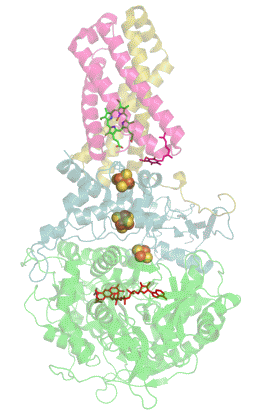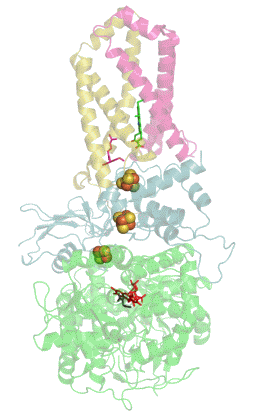
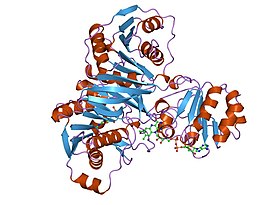
Сукцинатдегидрогеназа или сукцинат-убихинон-оксидорекдуктаза, также известная как комплекс II — белковый комплекс, расположенный во внутренней мембране митохондрий и мембранах многих прокариотических организмов. Одновременно участвует в цикле трикарбоновых кислот и дыхательной цепи переноса электронов.

Цитохром f — самая большая субъединица цитохром-b6/f комплекса шифр КФ 1.10.99.1. По своей структуре и функциям цитохром-b6/f комплекс аналогичен цитохром-b/c1 комплексу митохондрий и пурпурных бактерий. Цитохром f играет роль аналогичную цитохрому c1, несмотря на их различия по вторичной структуре.
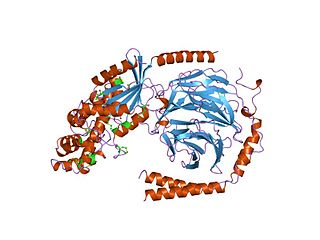
Гуанин-нуклеотид-связывающие белки — мембранно-связанные гетеротримерные белки, состоящие из трёх субъединиц, называемых «альфа», «бета» и «гамма». Гетеротримерные G-белки и связанные с ними G-белок-связанные рецепторы (GPCR) являются одной из самых распространённых внутриклеточных сигнальных систем в клетках животных, особенно млекопитающих. Они регулируют очень широкий круг процессов, таких, как сенсорные ощущения, рост, развитие, дифференцировку и размножение клеток, секрецию гормонов.

РНК-полимераза II — фермент эукариот, который катализирует транскрипцию ДНК, синтезирует предшественников мРНК и большинство мяРНК и микроРНК. Эта полимераза представляет собой комплекс массой 550 кДа, состоящий из 12 субъединиц. РНК-полимераза II является наиболее изученным типом РНК-полимеразы. Ей необходим широкий спектр транскрипционных факторов для того, чтобы связываться с генами выше промоторов и начинать транскрипцию.

Цитохро́м-b6f-ко́мплекс, или пластохинолпластоцианинредукта́за — мультибелковый комплекс, который осуществляет окисление пластохинолов и восстановление белка пластоцианина, обеспечивая, таким образом, транспорт электронов между реакционными центрами фотосистемы I (ФСI) и фотосистемы II (ФСII). Он восстанавливает маленький водорастворимый белок пластоцианин, который переносит электрон к ФСII. Аналогичную реакцию катализирует цитохром-bc1-комплекс электрон-транспортной цепи митохондрий. Цитохром-b6f-комплекс присутствует в тилакоидной мембране хлоропластов растений, водорослей и цианобактерий. Он функционально объединяет две фотосистемы в единую цепь переноса электронов от воды к НАДФ+, то есть является участником нециклического потока электронов. Кроме того, цитохромный комплекс вовлечён в циклический транспорт электронов, осуществляемый фотосистемой I.

Цитохро́м-bс1-ко́мплекс или убихинол-цитохром с-оксидоредуктаза, или комплекс III — мультибелковый комплекс дыхательной цепи переноса электронов и важнейший биохимический генератор протонного градиента на мембране митохондрий. Этот мультибелковый трансмембранный комплекс кодируется митохондриальным и ядерным геномами.

НАДН-дегидрогена́зный ко́мплекс, также называемый ко́мплекс I или НАДН-убихино́н-оксидоредукта́за — первый мультибелковый комплекс дыхательной цепи переноса электронов. Множество копий комплекса расположены в мембранах прокариотических организмов, способных к кислородному дыханию и внутренних мембранах митохондрий эукариотических клеток. По отношению к белкам человека комплекс I часто называют НАДН-дегидрогеназой.

ETF-дегидрогеназа или (электронпереносящий флавопротеин)-дегидрогеназа — фермент, который переносит электроны с электронпереносящего флавопротеина (ETF), растворённого в матриксе митохондрий, на мембранный переносчик электронов — убихинон. Этот фермент есть как у прокариот, так и эукариот. У этого энзима есть две простетические группы: флавин и железосерный кластер. Нехватка ETF-дегидрогеназы у человека вызывает наследственное заболевание глутаровая ацидемия второго типа. Кроме того, этот фермент — третья точка поступления электронов в ЭТЦ.

Фумаратредуктаза — фермент, катализирующий реакцию превращения фумарата в сукцинат. Является важным ферментом для метаболизма микроорганизмов, поскольку участвует в процессе анаэробного дыхания.
- Сукцинат + акцептор ↔ фумарат + восстановленный акцептор
Ацил-КоА-дегидрогеназы, также ACADs — семейство ферментов из класса оксидоредуктаз, которые катализируют реакции переноса протона (дегидрогенизация) от субстрата — ацил-КоА жирной кислоты на электрон-переносящий флавопротеин (FAD), участвуют в процессе β-окисления. Результатом реакции является образование двойной связи расположенной между атомами С2 (α) и С3 (β) в молекуле тиоэфира субстрата (ацил-КоА).

Ацил-КоА-дегидрогеназа жирных кислот со средней длиной цепи, также MCAD — один из митохондриальных ферментов семейства ацил-КоА-дегидрогеназ, класса оксидоредуктазы. Систематическое название фермента:электрон-переносящая флавопротеинзависимая 2,3-оксидорекдуктаза. Данный фермент участвует в процессе β-окисления жирных кислот. У человека ген, кодирующий данный фермент — ACADM, локализован на коротком плече (p-плече) 1-й хромосомы.
Ферредоксин-НАДФ+-редуктаза, сокращенно ФНР, фермент из класса оксидоредуктаз, катализирующий реакцию восстановления НАДФ+, используя в качестве донора электронов ферредоксин.
Карбоксидобактерии — группа аэробных хемолитоавтотрофных бактерий, получающих энергию при окислении угарного газа (СО), объединяемых на основании физиологических особенностей. Не являются таксономической группой. Включают α, β, и γ-протеобактерии, фирмикутов, и актинобактерий. Большинство известных карбоксидобактерий являются грамотрицательными бактериями.

Бифурка́ция электро́нов — это механизм разделения электронов в окислительно-восстановительной реакции: пара электронов соединения-донора разделяется на два отдельных электрона, которые переносятся на два одноэлектронных акцептора, при этом один электрон переносится на акцептор с более низким электрохимическим потенциалом за счёт энергии другого электрона, который переносится на акцептор с более высоким потенциалом, чем у исходного соединения.
Дегидрогеназный комплекс разветвлённых α-кетокислот, также дегидрогеназа α-кетокислот с разветвлённой цепью — мультиферментный комплекс, локализованный на внутренней мембране митохондрий. Данный комплекс катализирует реакцию окислительного декарбоксилирования разветвлённых α-кетокислот с короткой углеродной цепью. Дегидрогеназный комплекс структурно аналогичен пируватдегидрогеназному и α-кетоглутаратдегидрогеназному комплексам, образуя вместе с ними семейство митохондриальных ферментов, участвующих в окислительном декарбоксилировании α-кетокислот.

Перифери́ческие мембра́нные белки́, или вне́шние мембра́нные белки́, представляют собой мембранные белки, которые обычно непрочно и лишь временно прикрепляются к биологической мембране, с которой они связаны. Данные белки прикрепляются к интегральным мембранным белкам или проникают в периферические области липидного бислоя. Например, регуляторные белковые субъединицы многих ионных каналов и трансмембранных рецепторов можно определить как периферические мембранные белки. В отличие от интегральных мембранных белков, периферические мембранные белки обычно собираются в водорастворимый компонент, или фракцию, всех белков, выделенных в ходе процедуры очистки белков. Белки с GPI-якорями являются исключением из этого правила и могут иметь свойства очистки, аналогичные свойствам интегральных мембранных белков.