Эмбриональная индукция
Эмбриональная индукция — взаимодействие между частями развивающегося организма у многоклеточных, беспозвоночных и всех хордовых.
История
Первые упоминания о взаимодействии клеток зародышевых листков принадлежат открывшему их Х. Пандеру. В 1817 году Пандер показал, что зародышевые листки не образуют соответствующие им органы автономно. Скорее, каждый листок зародышевых клеток «пока не настолько независим, чтобы было ясно, чем он является, и по-прежнему нуждается в помощи своих „сестёр“ на пути развития; поэтому, несмотря на то, что им уже предначертан разный исход, все три листка сообща воздействуют друг на друга, пока каждый не достигает определённого уровня»[1].
Явление было открыто в 1901 году при изучении образования зачатка хрусталика глаз у зародышей земноводных. Гипотезу о механизме дифференцировки, получившем название эмбриональной индукции, на основании экспериментальных данных выдвинули Шпеман и Мангольд в 1924 году.
Гипотеза
Согласно этой гипотезе, существуют определённые клетки, которые действуют как организаторы на другие, подходящие для этого клетки. В условиях отсутствия клеток-организаторов такие клетки пойдут по другому пути развития, отличному от того, в котором они развивались бы в условиях присутствия организаторов. Проиллюстрировать это можно тем самым экспериментом 1924 года, показавшим, что дифференцировка в значительной степени контролируется влиянием клеток одного типа на клетки другого типа.
Эксперимент
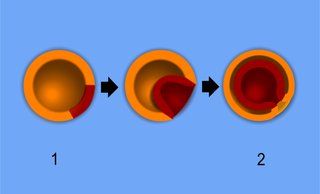
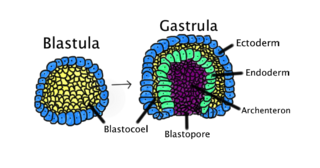
Г. Шпеман и его сотрудница Х. Мангольд открыли у зародышей амфибий «организатор». Контрольный эксперимент был проведён Хильдой Мангольд в 1921 году. Она вырезала кусочек ткани из дорсальной губы бластопора гаструлы гребенчатого тритона (Triturus cristatus) со слабопигментированными клетками и пересадила её в вентральную область другой гаструлы близкого вида, тритона обыкновенного (T. vulgaris), зародыш которого сильнее пигментирован. Эта естественная разница в пигментации позволила различить в химерном зародыше ткани донора и реципиента. Клетки дорсальной губы при нормальном развитии образуют хорду и мезодермальные сомиты (миотомы). После пересадки у гаструлы-реципиента из тканей трансплантата развивалась вторая хорда и миотомы. Над ними из эктодермы реципиента возникала новая дополнительная нервная трубка. В итоге это привело к образованию осевого комплекса органов второй личинки на том же зародыше.
Связь с общей концепцией регуляторных механизмов онтогенеза
Явление эмбриональной индукции тесно связано с такими понятиями, как морфоген и морфогенетическое поле (morphogenetic field). Ещё Шпеманом было показано, что инактивированные нагреванием ткани организатора сохраняют индуцирующую активность, и среда из-под изолированного организатора также индуцирует эктодерму.
Позже было показано, что многие ткани взрослых животных индуцируют нейрализацию эктодермы. Также были открыты вещества-индукторы, такие как хордин и ноггин (действуют косвенно, через подавление BMP (англ. bone morphogenetic protein) — эпидермального индуктора, его инактивация хордином и ноггином вызывает нейрализацию эктодермы), и многие другие.
Интересные факты
- Эмбриональная индукция — лишь один из механизмов онтогенеза. Многие процессы развития регулируются иными механизмами.
- Участок дорсальной губы бластопора, который при пересадке вызывает на новом месте образование мезодермы и нейроэктодермы получил название «организатор Шпемана».
- За своё открытие Ганс Шпеман получил в 1935 году Нобелевскую премию.
См. также
Примечания
- ↑ М. Дж. Ф, Барреси, С. Ф. Гилберт. Биология развития. — Москва: Лаборатория Знаний, 2022. — С. 37. — 800 с. — ISBN 978-5-00101-323-5.