Эндоспоры
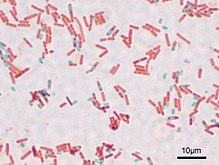
Эндоспо́ры — покоящиеся формы бактерий, которые образуются в результате скоординированной дифференцировки двух дочерних клеток, образовавшихся при удвоении генома исходной бактерии, причём одна из них проникает внутрь цитоплазмы другой дочерней клетки, которая становится внешней. Далее внешняя дочерняя клетка (её иногда называют спорангием) погибает программируемой гибелью, а внутренняя клетка (преспора) становится собственно эндоспорой и входит в состояние максимального физиологического покоя, при котором все физиологические процессы внутри споры останавливаются (гиперанабиоза). Она становится чрезвычайно резистентной к неблагоприятным условиям окружающей среды и может сохранять жизнеспособность в течение длительного времени[1].
Описаны два случая, когда материнская клетка даёт не одну, а две эндоспоры. Так, крупная (35 мкм длиной) некультивируемая бактерия Metabacterium polyspora, обитающая в желудочно-кишечном тракте некоторых грызунов, образует до девяти эндоспор на спорангий. Другая гигантская некультивируемая бактерия, Epulopiscium sp. (она может достигать размера 60 × 300 мкм), обитающая внутри тропической рыбы-единорога Naso tonganus[англ.], образует две эндоспоры в одном спорангии[2].
Спорогенные бактерии
Эндоспоры образуют представители около 25 родов из типа фирмикут: Bacillus, Clostridium, Dendrosporobacter[англ.], Desulfosporosinus[англ.], Desulfotomaculum[англ.], Halobacillus[англ.], Heliophilum, Oscillospira, Sporobacter, Sporobacterium[англ.], Sporohalobacter[англ.], Sporolactobacillus[англ.], Sporomusa[англ.], Sporosarcina[англ.], Sporotomaculum, Sulfobacillus[англ.], Thermoactinomyces и другие. В подавляющем большинстве случаев спорогенные бактерии являются грамположительными, хотя есть и исключения. Например, Sporomusa ovata[англ.] и Heliophilum fasciatum относятся к числу грамотрицательных бактерий[3].
Строение эндоспоры

Основные структурные элементы зрелой эндоспоры:
- цитоплазматический компартмент, или кор;
- комбинированная клеточная стенка, состоящая из внутренней стенки и наружной стенки (кортекса);
- цитоплазматическая мембрана эндоспоры (внутренняя мембрана);
- цитоплазматическая мембрана спорангия (внешняя мембрана);
- внутренний и внешний споровые покровы;
- наружный покров (экзоспорий)[4].

Расположение эндоспор внутри материнской клетки может быть различным, и тип спорообразования в некоторых случаях используют при идентификации неизвестной культуры. Иногда формирующаяся эндоспора достигает таких размеров, что расширяет спорангий посередине или с одного из концов[5].
Кор эндоспоры содержит хромосому и небольшие количества компонентов аппарата трансляции: рибосомы, тРНК, сопутствующие ферменты и белковые факторы. При этом нестабильные компоненты клеток, такие как мРНК и нуклеозидтрифосфаты, отсутствуют, но есть запасы стабильных предшественников АТФ — АДФ и АМФ[6].
Сложная система покровов эндоспоры делает её высокорезистентной к различным неблагоприятным факторам. Её белковые покровы перекрывают доступ к кортексу ферментам, разрушающим пептидогликан (например, лизоциму), а также защищают спору от агрессивных химических реагентов, таких как хлороформ. Мембраны спорангия и споры создают барьер проницаемости вокруг кора, не давая пройти к нему гидрофильным соединениям массой более 300 Да. Кроме того, низкое содержание воды в коре препятствует депуринизации геномной ДНК. Катионы двухвалентных металлов (прежде всего кальция) хелатируются молекулами дипиколиновой кислоты[англ.], которые синтезируются в спорангии, а потом доставляются в кор, что обеспечивает дополнительную защиту для ДНК. В коре эндоспор некоторых бактерий имеются особые белки, связывающиеся с ДНК и защищающие её не только от депуринизации, но и от разрушения сахарофосфатного остова. Наконец, прорастание эндоспоры сопровождается интенсивной репарацией повреждений в ДНК, накопленных во время периода покоя[7].
Споруляция

Цикл спорообразования протекает гораздо дольше (около 7 ч при 37 °С), чем деление с образованием других специализированных клеток. У Bacillus subtilis цикл споруляции включает семь стадий (0—VII), которые были предложены в 1960-х годах французским учёным Антуанетом Ритером.
- Стадия 0. Исходная вегетативная клетка.
- Стадия I. В настоящее время эту стадию не выделяют, так как характерных морфологических признаков она лишена, а мутанты, не способные её проходить, неизвестны.
- Стадия II подразделяется на три этапа. На этапе IIi происходит септирование: в материнской клетке образуется споровая септа и происходит асимметричное бинарное деление, при нём образуются протопласты разного размера. На этапах IIii—IIiii больший протопласт начинает поглощать меньший.
- Стадия III. Поглощение завершается, и меньший протопласт (преспора) образуется в цитоплазме большей клетки (спорангия).
- Стадия IV. Происходит синтез модифицированной клеточной стенки — кортекса, который окружает преспору.
- Стадия V. В цитоплазме спорангия откладываются белковые покровы будущей эндоспоры.
- Стадия VI. Стадия морфологически не выражена, во время неё происходит созревание эндоспоры. Она приобретает повышенную резистентность к факторам окружающей среды, входит в гиперанабиоз, становится способной к прорастанию.
- Стадия VII. Спорангий подвергается программируемой гибели и частично лизируется, из-за чего зрелая эндоспора выходит наружу[8].
Уникальная споровая септа закладывается при симметричном образовании двух Z-колец на ¼ и ¾ материнской клетки. Одно из них случайным образом выбирается при участии белка SpoIIA, второе разрушается. Далее в области Z-кольца, как при обычном делении, происходит кольцевая инвагинация мембраны, впоследствии заполняющаяся пептидогликаном. Далее он лизируется, и в результате сестринские компартменты оказываются разделёнными только двумя мембранами. Хромосома будущей эндоспоры, по последним данным, проникает в меньший компартмент после смыкания септы. Её перенос опосредует ДНК-транслоказа FtsK/SpoIIIE, формирующая в мембранах компартментов два сквозных кольцевых гексамерных канала, и C-концевые домены канальцевых белков снабжают процесс энергией АТФ. Процесс поглощения меньшего компартмента большим в целом соответствует эндоцитозу (который вообще бактериям не свойственен), его молекулярные механизмы изучены слабо[9].

В лабораторных условиях бактерии приступают к споруляции, когда в среде заканчивается легко метаболизируемый источник углерода, азота или фосфора, однако триггеры споруляции в естественной среде обитания бактерий неизвестны. Ключевое событие начала споруляции — фосфорилирование транскрипционного фактора Spo0A с помощью аутофосфорилирующих[англ.] протеинкиназ. Фосфорилирование Spo0A может запускаться тремя путями:
- Через киназу KinA, которая регистрирует окислительно-восстановительный и энергетический статус клетки.
- Через снижение уровня ГДФ и ГТФ в клетке, что свидетельствует о нехватке питательных веществ.
- Через белки, связанные с чувством кворума, то есть при достижении определённой плотности популяции бактерий[10].
В условиях, не благоприятствующих споруляции, фосфорилированный Spo0A дефосфорилируется фосфатазой Spo0E. Активный фосфорилированный Spo0A активирует транскрипцию регуляторов споруляции SpoIIA, SpoIIE и SpoIIG. Клетки, содержащие фосфорилированный Spo0A, выделяют белки, убивающие соседние клетки с неактивным Spo0A, чтобы использовать при споруляции их ресурсы. Также в запуске споруляции играет роль сигма-фактор σH, контролирующий гены spoIIID и spoIIIM, их продукты участвуют в образовании споровой септы. После формирования септы активность Spo0A возрастает в спорангии, но снижается в преспоре. Далее в преспоре активируется сигма-фактор σF, а в спорангии — σE. Также в преспоре синтезируется σG, который активируется только при завершении поглощения, а в спорангии на финальных стадиях спорообразования синтезируется сигма-фактор σK. Совместно сигма-факторы преспоры и спорангия активируют экспрессию генов, необходимых для формирования эндоспоры[11].
Состояние покоя и прорастание
Находящаяся в состоянии покоя эндоспора характеризуется гиперанабиозом и гиперрезистентностью. Она не проявляет метаболической активности, не содержит важнейших метаболитов, таких как АТФ и ацетил-CoA, находящиеся в ней ферменты неактивны. В состоянии гиперанабиоза споры могут сохранять жизнеспособность на протяжении огромного периода времени. Так, эндоспоры сибирской язвы в скотомогильниках сохраняют жизнеспособность в течение 500 лет, споры актиномицетов — до 7500 лет[12]. Имеются сведения, что споры Bacillus sp., добытые из кристаллов поваренной соли в Нью-Мексико, сохраняли жизнеспособность на протяжении 250 млн лет. Эндоспоры не погибают под действием высокой и низкой температуры, при высушивании, большом гидростатическом давлении, при УФ- и γ-излучении, под действием сильных окислителей, при повышенной кислотности и других неблагоприятных условиях[13]. Споры некоторых бактерий выдерживают даже кипячение в течение часа и более, поэтому растворы и инструменты стерилизуют в автоклавах с температурами стерилизации до 121 °C[12].
При благоприятных условиях эндоспора прорастает, то есть выходит из спорангия и превращается в нормальную вегетативную клетку. Индукторы прорастания могут быть как физиологическими (некоторые аминокислоты и сахара, пуриновые нуклеозиды и другие соединения или их смеси), так и нефизиологическими (минеральные соли, экзогенный дипиколинат кальция, лизоцим, катионные детергенты, сублетальный тепловой шок[англ.], давление от 100 до 600 МПа). Прорастание начинается через секунды после воздействия индуктора и далее от него не зависит. Сначала из эндоспоры выходят протоны, ионы калия, натрия и цинка, дипиколиновая кислота с хелатированными ионами Ca2+, входит вода. Далее с помощью специальных ферментов лизируется пептидогликан кортекса, продолжается регидратация, при которой кор набухает и стенка растягивается. Наконец возобновляется нормальный метаболизм и биосинтез макромолекул[14].
Визуализация
Эндоспоры хорошо просматриваются при помощи как светового, так и электронного микроскопа. Так как эндоспоры непроницаемы для многих красителей, их визуализируют как неокрашенные тельца на фоне остального прокрашенного содержимого бактериальной клетки. Существуют, однако, методы дифференциального окрашивания спор[англ.], с помощью которых споры становятся видны в световой микроскоп как синие тельца в розовой цитоплазме[12].
Примечания
- ↑ Пиневич, 2009, с. 257.
- ↑ Пиневич, 2009, с. 258—259.
- ↑ Пиневич, 2009, с. 258.
- ↑ Пиневич, 2009, с. 257—258.
- ↑ Нетрусов, Котова, 2012, с. 67—68.
- ↑ Нетрусов, Котова, 2012, с. 68—69.
- ↑ Пиневич, 2009, с. 261—262.
- ↑ Пиневич, 2009, с. 263.
- ↑ Пиневич, 2009, с. 263—264.
- ↑ Пиневич, 2009, с. 266—267.
- ↑ Пиневич, 2009, с. 267—268.
- ↑ 1 2 3 Нетрусов, Котова, 2012, с. 67.
- ↑ Пиневич, 2009, с. 259—261.
- ↑ Пиневич, 2009, с. 268—269.
Литература
- Пиневич А. В. Микробиология. Биология прокариотов: в 3 т. — СПб.: Издательство С.-Петербургского университета, 2009. — Т. III. — 457 с. — ISBN 978-5-288-04894-4.
- Нетрусов А. И., Котова И. Б. Микробиология. — 4-е изд., перераб. и доп. — М.: Издательский центр «Академия», 2012. — 384 с. — ISBN 978-5-7695-7979-0.
- Современная микробиология / Под ред. Й. Ленгелера, Г. Древса, Г. Шлегеля. — М.: Мир, 2005. — Т. 1. — 654 с.
| В библиографических каталогах |
|---|
| Патогенные бактерии | |
|---|---|
| Микрофлора человека |
|
| Субстратная специфичность | |
| Дыхание | |
| Генетика и размножение |
|
| Покоящиеся формы |
|
| См. также | |