
Гисто́ны — обширный класс ядерных белков, выполняющих две основные функции: они участвуют в упаковке нитей ДНК в ядре и в эпигенетической регуляции таких ядерных процессов, как транскрипция, репликация и репарация. Существует пять различных типов гистонов H1/Н5, H2A, H2B, H3, H4. Гистоны H2A, H2B, H3, H4, называемые кóровыми гистонами, формируют нуклеосому, представляющую собой белковую глобулу, вокруг которой накручена нить ДНК. Гистон H1/H5, называемый линкерным гистоном, связывается с внешней стороной нуклеосомы, фиксируя на ней нить ДНК. В хроматине гистоны составляют 25—40 % сухого веса. Благодаря высокому содержанию лизина и аргинина гистоны проявляют сильно оснóвные свойства. Гистоны непосредственно контактируют с ДНК и способны нейтрализовать отрицательный заряд фосфатных групп ДНК за счёт положительных зарядов аминокислотных остатков. Последовательность аминокислот в этих белках является консервативной и практически не различается в организмах различных таксонов. Гистоны присутствуют в ядрах эукариотических клеток; у бактерий гистонов нет, но они выявлены у архей группы Euryarchaea.

Карцинома — вид злокачественной опухоли, развивающейся из клеток эпителиальной ткани различных органов. По данным Всемирной организации здравоохранения, в 2018 году от этого заболевания умерли 9 600 000 человек.

Рили́н — белок, содержащийся в мозге и в других тканях и органах тела человека и других животных. Этот гликопротеин выполняет множество функций, важнейшей из которых является регулировка миграции и позиционирования нервных стволовых клеток в период фетального и раннего послеродового развития, необходимая для нормального формирования коры и других структур головного мозга. Во взрослом мозге рилин регулирует позиционирование нейронов, образуемых в процессе взрослого нейрогенеза, а также вносит вклад в работу механизмов памяти и обучения, модулируя синаптическую пластичность, усиливая и поддерживая долговременную потенциацию, стимулируя развитие дендритов и дендритных шипиков.
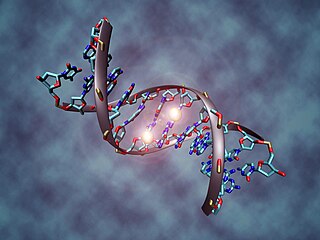
Метилирование ДНК — это модификация молекулы ДНК без изменения самой нуклеотидной последовательности ДНК, что можно рассматривать как часть эпигенетической составляющей генома.

Эпигенетическим наследованием называют наследуемые изменения в фенотипе или экспрессии генов, вызываемые механизмами, отличными от изменения последовательности ДНК. Такие изменения могут оставаться видимыми в течение нескольких клеточных поколений или даже нескольких поколений живых существ.
Белки группы polycomb — это семейство белков, которые способны ремоделировать хроматин. Эти белки-регуляторы были впервые описаны у дрозофил, где они подавляют гомеозисные гены, контролирующие индивидуальные отличия сегментов развивающегося эмбриона.

Эухромати́н, также активный или «открытый хромати́н» — участки хроматина, которые представляет собой неплотную и легко упакованную форму, обогащённую генами и часто находящуюся в состоянии активной транскрипции. Эухроматин отличается от гетерохроматина, который плотно упакован и менее доступен для транскрипции. Около 92 % генома человека является эухроматичным.

Антисмысловы́е РНК — одноцепочечные РНК, которые комплементарны мРНК, транскрибируемой в клетке, или гену-мишени. Механизмы действия антисмысловых РНК весьма разнообразны, они могут как подавлять, так и активировать экспрессию гена-мишени. Природные антисмысловые РНК есть и у прокариот, и у эукариот; они относятся к длинным некодирующим РНК как РНК длиной более 200 нуклеотидов. Синтетические антисмысловые РНК нашли широкое применение у исследователей в качестве инструмента для нокдауна генов. Антисмысловые РНК также находят медицинское применение.

Деацетилазы гистонов , — ферменты, катализирующие удаление ацетильной группы ε-N-ацетил-лизина гистонов, внесенные ферментами гистонацетилазами в остатки K3 и K14 гистона Н3 и K5, K8, K12 и K16 гистона Н4, а также остатки некоторых лизинов гистонов Н2А и Н2B. Модифицируя гистоны и изменяя конформацию хроматина, гистондеацетилазы играют важную роль в регуляции экспрессии генов. В то время как гиперацетилирование гистонов под действием гистонацетилаз обычно связано с повышением транскрипционной активности, гистондеацетилазы вызывают гипоацетилирование и вследствие, репрессию генов. Гипоацетилирование приводит к уменьшению промежутка между нуклеосомой и намотанной на неё ДНК. Более плотная упаковка ДНК уменьшает её доступность для транскрипционных факторов, что приводит к транскрипционной репрессии. Обычно гистондеацетилазы действуют в составе крупных комплексов, вместе с другими белками подавляющими активность хроматина. Субстратами гистондеацетилаз могут быть не только гистоны, но и некоторые другие белки.
Белки группы Trithorax — это белки, регулирующие структуру хроматина и участвующие в поддержании экспрессии генов. Это семейство белков гетерогенно, входящие в него белки образуют сложные белковые комплексы, способные связываться с хроматином. Так, функция самого белка Trithorax в составе комплекса TAC1 — поддержание локального открытого состояния хроматина путём монометилирования лизина-4 гистона H3 (H3K4me1), что сохраняет активность окрестных генов в ряду клеточных поколений. Действие этой гистоновой метки усиливается ацетилированием лизина-27 гистона H3 (H3K27ac), привносимой тем же комплексом TAC1. В дополнение к этому некоторые белки группы Trithorax ремоделируют хроматин, используя энергию АТФ для активации нуклеосом.
Эпигенетика — раздел генетики. Эпигенетика изучает наследуемые изменения активности генов во время роста и деления клеток — изменения синтеза белков, вызванных механизмами, не изменяющими последовательность нуклеотидов в ДНК. Эпигенетические изменения сохраняются в ряде митотических делений соматических клеток, а также могут передаваться следующим поколениям. Регуляторы синтеза белка — метилирование и деметилирование ДНК, ацетилирование и деацетилирование гистонов, фосфорилирование и дефосфорилирование транскрипционных факторов и другие внутриклеточные механизмы.
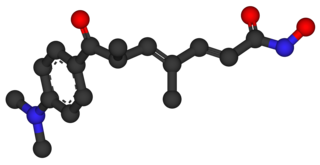
Трихостатин А — противогрибковый антибиотик, который является эффективным и специфическим ингибитором гистондезацетилазы (HDAC) клеток млекопитающих, как в естественных условиях, так и в пробирке. ТСА ингибирует клеточный цикл эукариотической клетки и индуцирует обратимую трансформацию клеток. Блокирует деление клетки в фазе G1.

ДНК (цитозин-5)-метилтрансфераза 3A — фермент, катализирующий перенос метильных групп к сайтам метилирования CpG в составе ДНК. Фермент кодируется у человека геном DNMT3A.

ДНК (цитозин-5)-метилтрансфераза 3-подобный фермент англ. DNA (cytosine-5)-methyltransferase 3-like, DNMT3L — фермент, кодируемый у человека геном DNMT3L.
Гистондеацетила́за 4 — белок, кодируемый у человека геном HDAC4, расположенным на 2-й хромосоме. Как и все ферменты группы гистондеацетилаз, близкой к сиртуинам, гистондеацетилаза 4 катализирует удаление ацетильных групп с остатков лизина в N-концевой части коровых гистонов, что изменяет структуру хроматина. Деацетилирование гистонов является одним из механизмов транскрипционной и эпигенетической регуляции, оказывает влияние на ход клеточного цикла и участвует в регуляции развития. Работа HDAC4 регулируется путём различных посттрансляционных модификаций и взаимодействий с разнообразными белками, иногда тканеспецифичными. Нарушение работы HDAC4 приводит к развитию многих заболеваний, в том числе раковых, поэтому ингибиторы HDAC4 могут иметь важное медицинское применение.

Гистондеацетилаза 10 — фермент, кодируемый у человека геном HDAC10 .
Парамута́ция — взаимодействие двух аллелей одного локуса, при котором один аллель вызывает наследуемые изменения в другом аллеле. Эти изменения могут заключаться в изменении паттерна метилирования ДНК или модификации гистонов. Аллель, индуцирующий эти изменения, называется парамутагенным, а тот аллель, который эпигенетически изменяется, называется парамутабельным. Парамутабельный аллель может иметь изменённые уровни экспрессии, которые могут сохраняться у потомства, унаследовавшего этот аллель, даже в отсутствие парамутагенного аллеля. Парамутации могут иметь место, например, у генетически идентичных растений, демонстрирующих совершенно разные фенотипы.
Эпигенети́ческие часы́ — это совокупность эпигенетических меток ДНК, позволяющая определить биологический возраст ткани, клетки или органа. Наиболее известным примером эпигенетических часов являются часы Стива Хорвата, учитыващие 353 эпигенетических маркера человеческого генома. Разработаны и другие версии эпигенетических часов: часы К. Вейднер, основанные на метилировании трёх CpG динуклеотидов, часы И. Флорат, сложные часы Г. Ханнума и часы К. Джулиани, показатель которых рассчитываются по метилированию трёх генов в образцах ДНК из дентина.
Эпигено́мика — раздел молекулярной биологии, изучающий совокупность эпигенетических модификаций генетического материала клетки с помощью высокопроизводительных методов. Эпигеномика аналогична геномике и протеомике, которые изучают геном и протеом клетки, соответственно.

Эпигенетика — это изучение изменений в экспрессии генов. Экспрессия происходит посредством метилирования ДНК, ацетилирования гистонов и модификации микро-РНК. Эпигенетические изменения подобного рода могут наследоваться и тогда они влияют на эволюцию. Современные исследования активно ведутся и уже понятно что эпигенетика оказывает большое влияние на все живые организмы.