
5-MeO-DMT, 5-метоксидиметилтриптамин — сильное психоактивное вещество из класса триптаминов. 5-MeO-DMT обнаруживается во многих растениях, а также в секрециях некоторых жаб. Является родственным таким веществам, как диметилтриптамин (DMT) и буфотенин (5-HO-DMT). Имеет тысячелетнюю историю использования в качестве энтеогена в шаманских практиках в Южной Америке.
Рецепторы, сопряжённые с G-белком,, также известные как семиспиральные рецепторы или серпентины, составляют большое семейство трансмембранных рецепторов. GPCR выполняют функцию активаторов внутриклеточных путей передачи сигнала, приводящими в итоге к клеточному ответу. Рецепторы этого семейства обнаружены только в клетках эукариот: у дрожжей, растений, хоанофлагеллят и животных. Эндогенные лиганды-агонисты, которые связываются и активируют эти рецепторы, включают гормоны, нейромедиаторы, светочувствительные вещества, пахучие вещества, феромоны и варьируются в своих размерах от небольших молекул и пептидов до белков. Нарушение работы GPCR приводит к возникновению множества различных заболеваний, а сами рецепторы являются мишенью до 40 % выпускаемых лекарств. Точный размер надсемейства GPCR не известен, но почти 800 различных человеческих генов были предсказаны из анализа последовательности генома. Несмотря на многочисленные схемы, было предложено разделить надсемейство на три основных класса.
Дофаминовые рецепторы — класс трансмембранных метаботропных G-белок-связанных клеточных рецепторов, играющих важную роль в функционировании центральной нервной системы позвоночных. Основной эндогенный лиганд-агонист этих рецепторов — дофамин. Дофаминовые рецепторы участвуют в процессах мотивации, обучения, тонкой моторной координации, модулирования нейроэндокринных сигналов. Этот класс включает пять типов рецепторов: D1, D2, D3, D4 и D5.

Серотониновые рецепторы (5-HT-рецепторы) — мембранные рецепторы 5-гидрокситриптамина (5-HT), нейромедиатора и гормона, известного под названием серотонин, взаимодействующие также с множеством медицинских препаратов и психоактивных веществ. Активация рецепторов запускает внутриклеточные процессы, влияющие на активность других медиаторных систем — глутаматной, дофаминовой и ГАМК.
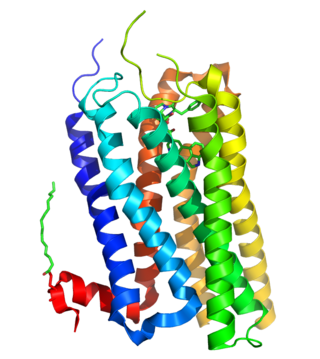
5-HT2A-рецептор у млекопитающих является одним из подтипов 5-HT2-рецепторов, подсемейства серотониновых рецепторов. Он является метаботропным G-белок-сопряжённым рецептором. Рецептор этого подтипа (5-HT2A) является основным возбуждающим подтипом рецепторов среди всех G-белок-сопряжённых подтипов рецепторов для серотонина (5-HT). Однако рецепторы подтипа 5-HT2A способны, наоборот, оказывать ингибирующее, тормозящее воздействие в некоторых областях мозга, таких, как зрительная кора и орбитофронтальная кора.
5-HT1A-рецептор — подтип серотониновых рецепторов подсемейства 5-HT1-рецепторов. Эндогенным лигандом-агонистом для них является моноаминовый нейромедиатор серотонин (5-гидрокситриптамин, 5-HT). Рецепторы этого подтипа относятся к семейству трансмембранных метаботропных G-белок-связанных рецепторов и связаны с гетеротримерным ингибиторным G-белком, так называемым Gi / Go. Этот подтип рецепторов опосредует тормозящую нейротрансмиссию. Ген, кодирующий белок этого рецептора у человека, обозначается HTR1A.
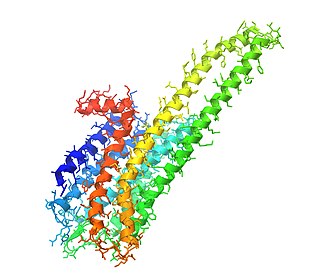
5-HT2C-рецептор — подтип серотониновых рецепторов подсемейства 5-HT2-рецепторов, который связывается с эндогенным нейромедиатором серотонином (5-гидрокситриптамином, 5-HT). Это метаботропный G-белок-сопряжённый рецептор, который связан с альфа-субъединицей белка Gq/G11 и медиирует возбуждающую нейротрансмиссию. Человеческий ген белка этого рецептора называется HTR2C. У человека он находится в хромосоме Х. Поскольку у мужчин имеется всего одна копия этого гена, а у женщин две, причём одна из копий этого гена репрессируется (и в разных клетках репрессированными могут оказаться разные аллели этого гена), то полиморфизмы этого гена влияют на поведенческие характеристики и на предрасположенность к тем или иным заболеваниям в разной степени у мужчин и у женщин.
5-HT2B-рецептор — белок одного из подтипов серотониновых рецепторов подсемейства 5-HT2-рецепторов. У человека этот рецепторный белок кодируется геном HTR2B. Рецепторы подтипа 5-HT2B относятся к семейству метаботропных G-белок-связанных рецепторов, и служат для связывания нейромедиатора серотонина.

5-HT₆-рецептор — подтип серотониновых рецепторов, который связывает эндогенный нейромедиатор серотонин. Он относится к классу G-белок-связанных метаботропных рецепторов и связан с Gsα. Он медиирует возбуждающую нейротрансмиссию. Этот рецепторный белок у человека кодируется геном HTR6.

5-HT₇-рецептор — это один из подтипов серотониновых рецепторов, относящийся к семейству метаботропных G-белок-связанных рецепторов на мембране клетки. Он активируется нейромедиатором серотонином. Рецепторы подтипа 5-HT₇ связаны с гетеротримерным G-белком Gs, который стимулирует образование внутриклеточных сигнальных молекул — молекул цАМФ. Он экспрессируется в различных тканях человека, в особенности в мозге, в желудочно-кишечном тракте и в кровеносных сосудах. В последнее время 5-HT₇-рецептор является мишенью для разработки новых лекарств, предназначенных для лечения различных заболеваний, в частности, лекарств для лечения депрессий, то есть антидепрессантов. Рецепторы подтипа 5-HT₇ кодируются геном HTR7, который у человека транскрипционируется путём альтернативного сплайсинга в 3 различных сплайс-варианта — 5-HT₇(a), 5-HT₇(b), 5-HT₇(d). Вариант 5-HT₇(c) у человека отсутствует, зато имеется у крыс, у которых отсутствует 5-HT₇(d).

5-HT5A-рецептор — один из подтипов серотониновых рецепторов. Белок этого рецептора у человека кодируется геном HTR5A. Это метаботропный G-белок-связанный рецептор, для которого нейромедиатором служит серотонин.

5-HT₄-рецептор — это один из подтипов серотониновых рецепторов. Он является метаботропным G-белок-связанным рецептором. Его стимуляция повышает продукцию циклического АМФ в клетке. Его эндогенным лигандом, как и для других типов серотониновых рецепторов, является нейромедиатор серотонин. У человека этот рецепторный белок кодируется геном HTR4. Продуктом этого гена является гликозилированный трансмембранный рецепторный белок, который обнаруживается как в периферических тканях, так и в центральной нервной системе и который модулирует и регулирует высвобождение различных нейромедиаторов. Этот белок имеет несколько вариантов транскрипции (изоформ) с различными C-терминальными окончаниями, однако по состоянию на начало 2014 года ещё не все варианты транскрипции данного гена полностью изучены и охарактеризованы.

Моноаминовые нейромедиаторы — это нейромедиаторы и нейромодуляторы, которые содержат одну аминогруппу, соединённую с ароматическим кольцом цепью из двух атомов углерода (-CH2-CH2-). Все моноаминовые нейромедиаторы являются производными ароматических аминокислот, таких, как фенилаланин, тирозин, триптофан, из которых они образуются при помощи ферментов, называемых декарбоксилазами ароматических аминокислот (в частности, триптофан-декарбоксилаза). Моноаминергические системы, то есть нервные сети, нейроны которых используют моноамины для обеспечения нейротрансмиссии, вовлечены в регуляцию многих процессов, в частности эмоций, когнитивных процессов, памяти, внимания, возбуждения и др. Также моноаминовые нейромедиаторы играют важную роль в обеспечении продукции и секреции нейротрофинов, в частности нейротрофина-3 астроцитами, что важно для обеспечения целостности нейронов, их нормальной дифференцировки и развития, трофической поддержки и для обеспечения их устойчивости к апоптозу. Лекарства, которые повышают или понижают активность тех или иных моноаминовых нейромедиаторов, могут быть использованы для фармакотерапии больных с теми или иными психическими расстройствами, такими, как депрессия, тревожные состояния, шизофрения.
Серотониновые рецепторы подтипа 5-HT1A играют в организме человека и животных множество очень важных физиологических ролей. Они являются наиболее распространённым в мозгу и периферических тканях подтипом серотониновых рецепторов, что подчёркивает их важную физиологическую роль.
Антагонистами 5-HT1A подтипа серотониновых рецепторов являются многие химические вещества и лекарства, в частности, некоторые бета-блокаторы, некоторые типичные и атипичные антипсихотики, некоторые антимигренозные препараты.
Рецепторы подтипа 5-HT₁A вовлечены в нейромодуляцию. В частности, активация этих рецепторов агонистами приводит к снижению артериального давления и урежению частоты сердечных сокращений благодаря центральным механизмам действия, а также благодаря индуцированию периферической вазодилатации и за счёт стимуляции активности блуждающего нерва. Эти эффекты являются результатом активации 5-HT₁A-рецепторов, находящихся в сосудодвигательном (прессорном) центре продолговатого мозга, в его рострально-вентролатеральной части. Симпатолитическое гипотензивное лекарство урапидил является не только антагонистом α₁-адренорецепторов и агонистом α₂-адренорецепторов, но и агонистом 5-HT₁A-рецепторов. И было показано, что его свойства как агониста 5-HT₁A-рецепторов привносят свой вклад в его гипотензивный эффект. Вазодилатация поверхностных сосудов кожи через активацию центральных 5-HT₁A-рецепторов увеличивает теплоотдачу с поверхности тела и тем самым способствует снижению температуры тела.
Активация 5-HT₁A-рецепторов вызывает секрецию различных гормонов, включая кортизол, АКТГ, окситоцин, пролактин, соматотропин, β-эндорфин. В отличие от 5-HT2-рецепторов, активация 5-HT₁A-рецепторов не оказывает влияния на секрецию вазопрессина или ренина. Предполагается, что секреция окситоцина может привносить свой вклад в просоциальные, антиагрессивные, анксиолитические эффекты, наблюдаемые при активации 5-HT₁A-рецепторов. Секреция β-эндорфина может привносить свой вклад в антидепрессивный, анксиолитический и анальгетический эффекты.

Ксиламидин — лекарство, которое действует как антагонист серотониновых рецепторов подтипа 5HT2A и, в меньшей степени, 5-HT1A. Ксиламидин не преодолевает ГЭБ, что делает его удобным для блокирования периферических серотонинергических реакций и желудочно-кишечного тракта, без вызывания таких центральных эффектов блокады 5-HT2A, как сонливость и седация, или снижения эффективности одновременно применяемых центральных 5-HT2A-агонистов.

Лиганд-зависимые ионные каналы, лиганд-управляемые или лиганд-активируемые ионные каналы — также обычно называемые ионотропными рецепторами, представляют собой группу трансмембранных белков ионных каналов, которые позволяют ионам, например, Na+, K+, Ca2+ и/или Cl-, проходить через биологическую мембрану, посредством изменения конформации (открытия) в ответ на связывание химического мессенджера (то есть лиганда), такого как, например, молекулы нейромедиатора.
5-HT3-рецептор, сокр. 5-HT3 — подтип серотониновых рецепторов, который относится к суперсемейству цис-петлевых лиганд-зависимых ионных каналов (ионотропных рецепторов) и поэтому структурно и функционально отличается от всех других рецепторов 5-HT-рецепторов (5-гидрокситриптамина или серотонина), которые являются рецепторами, сопряжённые с G-белком (GPCR). 5-HT3 представляет собой селективный катионный канал, он обеспечивает деполяризацию и возбуждение нейронов в центральной и периферической нервной системе.