5-HT3
5-HT3-рецептор, сокр. 5-HT3 — подтип серотониновых рецепторов, который относится к суперсемейству цис-петлевых лиганд-зависимых ионных каналов (ионотропных рецепторов) и поэтому структурно и функционально отличается от всех других рецепторов 5-HT-рецепторов (5-гидрокситриптамина или серотонина), которые являются рецепторами, сопряжённые с G-белком (GPCR)[1][2][3]. 5-HT3 представляет собой селективный катионный канал, он обеспечивает деполяризацию и возбуждение нейронов в центральной и периферической нервной системе[1].
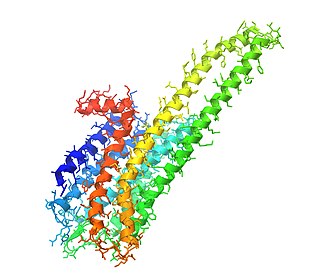
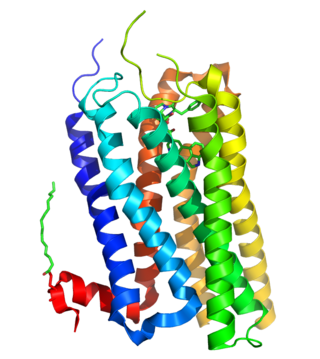
Как и в случае других лиганд-управляемых ионных каналов, рецептор 5-HT3 состоит из пяти субъединиц, расположенных вокруг центральной ионопроводящей поры, которая проницаема для ионов натрия (Na+), калия (K+) и кальция (Ca2+). Связывание нейромедиатора 5-гидрокситриптамина (серотонина) с рецептором 5-HT3 открывает канал, что, в свою очередь, приводит к возбуждающему ответу в нейронах (потенциалу действия). 5-HT3-рецепторы имеют незначительную проницаемость для анионов[1]. По структуре они наиболее гомологичны с никотиновыми рецепторами ацетилхолина.
История открытия
Идентификация 5-НТ3-рецептора не проводилась до 1986 года, в связи с отсутствием селективных фармакологических средств[4]. Однако, как только было установлено, что данный рецептор играет заметную роль в рвоте, вызванной химиотерапией и радиотерапией, и сопутствующее развитие селективных антагонистов рецептора 5-HT3 для подавления этих побочных эффектов вызвало большой интерес со стороны фармацевтической промышленности[2][5] и, как следствие, быстро последовала идентификация 5-HT3-рецепторов в клеточных линиях и нативных тканях[4].
Структура
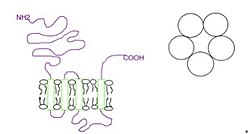
Функционально ионный канал может состоять из пяти идентичных субъединиц 5-HT3A (гомопентамерных) или в совокупности из 5-HT3A и одной из четырёх других (гетеропентамерных) субъединиц — 5-HT3B[6][7][8][9], 5-HT3C, 5-HT3D или 5-HT3E[10]. Похоже, что только субъединицы 5-HT3A образуют функциональные гомопентамерные каналы. Все другие подтипы субъединиц должны гетеропентамеризоваться с субъединицами 5-HT3A, чтобы сформировать функциональные каналы. Кроме того, в настоящее время не обнаружено каких-либо фармакологических различий между гетеромерными 5-HT3AC, 5-HT3AD, 5-HT3AE и гомомерным рецептором 5-HT3A[11]. N-концевое гликозилирование рецепторных субъединиц является критическим для сборки субъединиц и переноса их к плазматической мембране нейронов[12]. Субъединицы окружают центральный ионный канал псевдосимметричным образом (рисунок 1). Каждая субъединица содержит внеклеточный N-концевой домен, который содержит ортостерический сайт связывания лиганда; трансмембранный домен, состоящий из четырёх взаимосвязанных альфа-спиралей (M1-M4) с внеклеточной петлей M2-M3, вовлеченной в механизм стробирования; большой цитоплазматический домен между спиралями М3 и М4, участвующий в перемещении и регуляции рецепторов; и короткий внеклеточный С-конец (рисунок 1)[1]. В то время как внеклеточный домен является сайтом действия агонистов и конкурентных антагонистов, трансмембранный домен содержит центральную ионную пору, рецепторный затвор и селективный фильтр, который позволяет ионам пересекать клеточную мембрану[2].
Ген
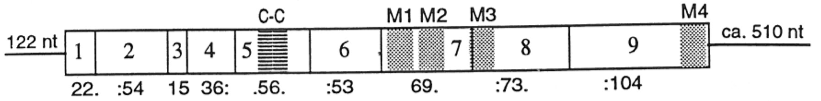
Ген 5-HT3-рецептора человека локализован в 11-ой хромосоме (локус 11q23.1-q23.2). Структурно сходен с геном мыши, который имеет 9 экзонов, размер ~ 13 кб. Четыре из его интронов находятся точно в том же положении, что и интроны в гомологичном гене α7-ацетилхолинового рецептора, что чётко подтверждает их эволюционную связь[13][14]. Также были идентифицированы дополнительные гены, которые кодируют субъединицы 5-HT3-рецептора. HTR3A и HTR3B гены, кодирующие соответственно субъединицы 5-HT3A и 5-HT3B и, кроме того, гены HTR3C, HTR3D и HTR3E, кодирующие субъединицы 5-HT3C, 5-HT3D и 5-HT3E. Гены HTR3C и HTR3E, по-видимому, при экспрессии не образуют функциональных гомомерных каналов, однако, при совместной экспрессии с HTR3A они образуют гетеромерный комплекс с пониженной или повышенной эффективностью серотонина. Патофизиологическая роль данных дополнительных субъединиц ещё не определена[15].

Экспрессия. Гены, кодирующие 5-HT3C, 5-HT3D и 5-HT3E субъединицы, имеют тенденцию к периферическому ограниченному типу экспрессии с высоким уровнем в кишечнике. Например, в двенадцатиперстной кишке и желудке человека количество мРНК 5-HT3C и 5-HT3E может быть больше, чем 5-HT3A и 5-HT3B.
Полиморфизм. У пациентов, получающих химиотерапевтические препараты, определённый полиморфизм гена HTR3B может предсказать успешное антиэметическое (противорвотное) лечение. Это может указывать на то, что 5-HTR3B субъединицу рецептора возможно использовать в качестве биомаркера эффективности противорвотных препаратов.
Распределение в организме
5-HT3-рецептор экспрессируется по всей центральной и периферической нервной системе и опосредует различные физиологические функции[4]. На клеточном уровне было показано, что постсинаптические 5-HT3-рецепторы опосредуют быструю возбуждающую синаптическую передачу в неокортикальных интернейронах крыс, миндалине и гиппокампе, а также в зрительной коре хорька[16][17][18][19] . 5-HT3-рецепторы также присутствуют на пресинаптических нервных окончаниях. Существуют также неубедительные доказательства участия данных рецепторов в модуляции высвобождения нейромедиаторов[20][21][22].
Проявляемые эффекты
При активации рецептора агонистами происходит открытие катионного канала, что приводит к следующим эффектам:
- В ЦНС: возбуждение центра тошноты и рвоты в стволе головного мозга, тревожность[23], склонность к судорогам[24], преноцицепция[25][26].
- В ПНС: возбуждение нейронов (происходит в вегетативных, ноцицептивных нейронах), рвота[23].
Эффекторы
К эффекторам 5-HT3-рецепторов относятся биологически активные молекулы — лиганды, способные влиять на функции данных белков, активируя или ингибируя их.
Агонисты
Агонисты рецептора включают в себя:
- Цереулид
- 2-метил-5-НT
- Альфа-метилтриптамин
- Буфотенин
- Хлорофенилбигуанид[23]
- Этиловый спирт
- Ибогаин
- Фенилбигуанид
- Квипазин
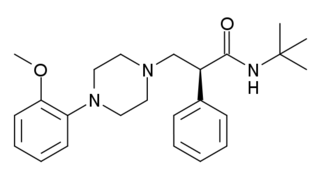
- RS-56812: мощный и селективный частичный агонист 5-HT3, селективность в 1000 раз выше по сравнению с другими рецепторами серотонина
- SR-57227
- Варениклин[27]
- YM-31636[28]
Антагонисты
Антагонисты рецептора (отсортированные по их соответствующему терапевтическому применению) включают:
Гастрокинетики:
- Алосетрон
- Батаноприд
- Метоклопрамид (антагонизм проявляется в высоких дозах)
- Рензаприд
- Закоприд
- М1, основной активный метаболит мосаприда
Другие:
- 3-тропанил-индол-3-карбоксилат
- Ламотриджин (средство против эпилепсии и биполярного расстройства)
- Мемантин (средство для лечения пациентов со средней и тяжёлой степенью болезни Альцгеймера)
- Ментол[29]
- Туйон
Положительные аллостерические модуляторы
Аллостерические модуляторы не являются агонистами рецептора, однако, они увеличивают сродство или эффективность рецепторов к молекулам эффектора (агониста), посредством активации аллостерической области, отдалённой от сайтов связывания лигандов:
Производные индола:
- 5-Хлориндол[30]
Органические анестетики с малой молекулярной массой:
См. также
Примечания
- ↑ 1 2 3 4 Barnes N. M., Hales T. G., Lummis S. C., Peters J. A. The 5-HT3 receptor--the relationship between structure and function (англ.) // Neuropharmacology : journal. — 2009. — January (vol. 56, no. 1). — P. 273—284. — doi:10.1016/j.neuropharm.2008.08.003. — PMID 18761359. — PMC 6485434.
- ↑ 1 2 3 Thompson A. J., Lummis S. C. 5-HT3 Receptors (англ.) // Current Pharmaceutical Design[англ.] : journal. — 2006. — Vol. 12, no. 28. — P. 3615—3630. — doi:10.2174/138161206778522029. — PMID 17073663. — PMC 2664614.
- ↑ Reeves D. C., Lummis S. C. The molecular basis of the structure and function of the 5-HT3 receptor: a model ligand-gated ion channel (review) (англ.) // Molecular Membrane Biology[англ.] : journal. — 2002. — Vol. 19, no. 1. — P. 11—26. — doi:10.1080/09687680110110048. — PMID 11989819.
- ↑ 1 2 3 Yakel, J. L. The 5-HT3 receptor channel: function, activation and regulation in Pharmacology of Ionic Channel Function: Activators and Inhibitors (Handbook of Experimental Pharmacology) (англ.) / Endo, M; Kurachi, Y; Mishina, M.. — Berlin: Springer-Verlag, 2000. — Vol. 147. — P. 541—560. — ISBN 3-540-66127-1.
- ↑ Thompson A. J., Lummis S. C. The 5-HT3 receptor as a therapeutic target (англ.) // Expert Opin Ther Targets[англ.] : journal. — 2007. — Vol. 11, no. 4. — P. 527—540. — doi:10.1517/14728222.11.4.527. — PMID 17373882. — PMC 1994432.
- ↑ Davies P. A., Pistis M., Hanna M. C., Peters J. A., Lambert J. J., Hales T. G., Kirkness E. F. The 5-HT3B subunit is a major determinant of serotonin-receptor function (англ.) // Nature : journal. — 1999. — Vol. 397, no. 6717. — P. 359—363. — doi:10.1038/16941. — . — PMID 9950429.
- ↑ Dubin A. E., Huvar R., D'Andrea M. R., Pyati J., Zhu J. Y., Joy K. C., Wilson S. J., Galindo J. E., Glass C. A., Luo L., Jackson M. R., Lovenberg T. W., Erlander M. G. The pharmacological and functional characteristics of the serotonin 5-HT3A receptor are specifically modified by a 5-HT3B receptor subunit (англ.) // J Biol Chem : journal. — 1999. — Vol. 274, no. 43. — P. 30799—30810. — doi:10.1074/jbc.274.43.30799. — PMID 10521471.
- ↑ Monk S. A., Desai K., Brady C. A., Williams J. M., Lin L., Princivalle A., Hope A. G., Barnes N. M. Generation of a selective 5-HT3B subunit-recognising polyclonal antibody; identification of immunoreactive cells in rat hippocampus (англ.) // Neuropharmacology : journal. — 2001. — Vol. 41, no. 8. — P. 1013—1016. — doi:10.1016/S0028-3908(01)00153-8. — PMID 11747906.
- ↑ Boyd G. W., Low P., Dunlop J. I., Ward M., Vardy A. W., Lambert J. J., Peters J., Conolly C. N. Assembly and cell surface expression of homomeric and heteromeric 5-HT3 receptors: The role of oligomerisation and chaperone proteins (англ.) // Mol Cell Neurosci[англ.] : journal. — 2002. — Vol. 21, no. 1. — P. 38—50. — doi:10.1006/mcne.2002.1160. — PMID 12359150.
- ↑ Niesler B., Walstab J., Combrink S., Moeller D., Kapeller J., Rietdorf J., Boenisch H., Goethert M., Rappold G., Bruess M. Characterization of the Novel Human Serotonin Receptor Subunits 5-HT3C, 5- HT3D and 5-HT3E (англ.) // Mol Pharmacol[англ.] : journal. — 2007. — Vol. 72, no. Mar 28. — P. 8—17. — doi:10.1124/mol.106.032144. — PMID 17392525.
- ↑ Niesler, Beate. 5-HT3 receptors: potential of individual isoforms for personalised therapy (англ.) // Current Opinion in Pharmacology : journal. — Elsevier, 2011. — February (vol. 11, no. 1). — P. 81—86. — doi:10.1016/j.coph.2011.01.011. — PMID 21345729.
- ↑ Quirk, Phillip L.; Rao, Suma; Roth, Bryan L.; Siegel, Ruth E. Three putative N-glycosylation sites within the murine 5-HT3A receptor sequence affect plasma membrane targeting, ligand binding, and calcium influx in heterologous mammalian cells (англ.) // Journal of Neuroscience Research[англ.] : journal. — 2004. — 15 August (vol. 77, no. 4). — P. 498—506. — ISSN 0360-4012. — doi:10.1002/jnr.20185. — PMID 15264219.
- ↑ 1 2 3 Uetz, P; Abdelatty, F; Villarroel, A; Rappold, G; Weiss, B; Koenen, M. Organisation of the murine 5-HT3 receptor gene and assignment to human chromosome 11 (англ.) // FEBS Letters[англ.] : journal. — 1994. — Vol. 339, no. 3. — P. 302—306. — doi:10.1016/0014-5793(94)80435-4. — PMID 8112471.
- ↑ Uetz, P. (1992) Das 5HT3-Rezeptorgen der Maus. Diploma Thesis, University of Heidelberg, 143 pp.
- ↑ Sanger G. J. 5-hydroxytryptamine and the gastrointestinal tract: where next? (англ.) // Trends in Pharmacological Sciences[англ.] : journal. — Cell Press, 2008. — September (vol. 29, no. 9). — P. 465—471. — doi:10.1016/j.tips.2008.06.008. — PMID 19086255.
- ↑ Férézou I., Cauli B., Hill E. L., Rossier J., Hamel E., Lambolez B. 5-HT3 receptors mediate serotonergic fast synaptic excitation of neocortical vasoactive intestinal peptide/cholecystokinin interneurons (англ.) // J Neurosci[англ.] : journal. — 2002. — Vol. 22, no. 17. — P. 7389—7397. — doi:10.1523/JNEUROSCI.22-17-07389.2002. — PMID 12196560.
- ↑ Kazuyoshi Kawa. Distribution and Functional Properties of 5HT3 Receptors in the Rat Hippocampus Dentate Gyrus (англ.) // Journal of Neurophysiology[англ.] : journal. — 1994. — Vol. 71, no. 5. — P. 1935—1947. — doi:10.1152/jn.1994.71.5.1935. — PMID 7520482.
- ↑ Sugita S., Shen K. Z., North R. A. 5-hydroxytryptamine is a fast excitatory transmitter at 5-HT3 receptors in rat amygdala (англ.) // Neuron[англ.] : journal. — Cell Press, 1992. — Vol. 8, no. 1. — P. 199—203. — doi:10.1016/0896-6273(92)90121-S. — PMID 1346089.
- ↑ Roerig B., Nelson D. A., Katz L. C. Fast synaptic signaling by nicotinic acetylcholine and serotonin 5-HT3 receptors in developing visual cortex (англ.) // J Neurosci[англ.] : journal. — 1992. — Vol. 17, no. 21. — P. 199—203. — PMID 9334409.
- ↑ van Hooft J. A., Vijverberg H. P. 5-HT3 receptors and neurotransmitter release in the CNS: a nerve ending story? (англ.) // Trends Neurosci[англ.] : journal. — 2000. — Vol. 23, no. 12. — P. 605—610. — doi:10.1016/S0166-2236(00)01662-3. — PMID 11137150.
- ↑ Rondé P., Nichols R. A. High calcium permeability of serotonin 5-HT3 receptors on presynaptic nerve terminals from rat striatum (англ.) // J Neurochem[англ.] : journal. — 1998. — Vol. 70, no. 3. — P. 1094—1103. — doi:10.1046/j.1471-4159.1998.70031094.x. — PMID 9489730.
- ↑ Rondé P., Nichols R. A. 5-HT3 receptors induce rises in cytosolic and nuclear calcium in NG108-15 cells via calcium-induced calcium release (англ.) // Cell Calcium[англ.] : journal. — 1997. — Vol. 22, no. 5. — P. 357—365. — doi:10.1016/S0143-4160(97)90020-8. — PMID 9448942.
- ↑ 1 2 3 4 5 Rang, H. P. Pharmacology (англ.). — Edinburgh: Churchill Livingstone[англ.], 2003. — ISBN 0-443-07145-4., page 187.
- ↑ Gholipour T., Ghasemi M., Riazi K., Ghaffarpour M., Dehpour A. R. Seizure susceptibility alteration through 5-HT(3) receptor: modulation by nitric oxide (англ.) // Seizure : journal. — 2010. — January (vol. 19, no. 1). — P. 17—22. — doi:10.1016/j.seizure.2009.10.006. — PMID 19942458.
- ↑ Patel, Ryan; Dickenson, Anthony H. Modality selective roles of pro-nociceptive spinal 5-HT2A and 5-HT3 receptors in normal and neuropathic states (англ.) // Neuropharmacology : journal. — 2018. — September (vol. 143). — P. 29—37. — ISSN 0028-3908. — doi:10.1016/j.neuropharm.2018.09.028. — PMID 30240783. — PMC 6277848.
- ↑ Suzuki, Rie; Rahman, Wahida; Rygh, Lars J; Webber, Mark; Hunt, Stephen P; Dickenson, Anthony H. Spinal-supraspinal serotonergic circuits regulating neuropathic pain and its treatment with gabapentin (англ.) // Pain : journal. — 2005. — October (vol. 117, no. 3). — P. 292—303. — ISSN 0304-3959. — doi:10.1016/j.pain.2005.06.015. — PMID 16150546.
- ↑ Mineur Y. S., Picciotto M. R. Nicotine receptors and depression: revisiting and revising the cholinergic hypothesis (англ.) // Trends Pharmacol. Sci.[англ.] : journal. — 2010. — December (vol. 31, no. 12). — P. 580—586. — doi:10.1016/j.tips.2010.09.004. — PMID 20965579. — PMC 2991594.
- ↑ Imanishi, N.; Iwaoka, K.; Koshio, H.; Nagashima, S. Y.; Kazuta, K. I.; Ohta, M.; Sakamoto, S.; Ito, H.; Akuzawa, S.; Kiso, T.; Tsukamoto, S. I.; Mase, T. New thiazole derivatives as potent and selective 5-hydroxytriptamine 3 (5-HT3) receptor agonists for the treatment of constipation (англ.) // Bioorganic & Medicinal Chemistry[англ.] : journal. — 2003. — Vol. 11, no. 7. — P. 1493—1502. — doi:10.1016/S0968-0896(02)00557-6.
- ↑ Ashoor, A.; Nordman, J.; Veltri, D.; Susan Yang, K. -H.; Shuba, Y.; Al Kury, L.; Sadek, B.; Howarth, F. C.; Shehu, A.; Kabbani, N.; Oz, M. Menthol Inhibits 5-Ht3 Receptor-Mediated Currents (англ.) // Journal of Pharmacology and Experimental Therapeutics[англ.] : journal. — 2013. — Vol. 347, no. 2. — P. 398—409. — doi:10.1124/jpet.113.203976. — PMID 23965380.
- ↑ Newman, A. S.; Batis, N; Grafton, G; Caputo, F; Brady, C. A.; Lambert, J. J.; Peters, J. A.; Gordon, J; Brain, K. L.; Powell, A. D.; Barnes, N. M. 5-Chloroindole: A potent allosteric modulator of the 5-HT3 receptor (англ.) // British Journal of Pharmacology[англ.] : journal. — 2013. — Vol. 169, no. 6. — P. 1228—1238. — doi:10.1111/bph.12213. — PMID 23594147. — PMC 3831704.
- ↑ Davies P. A. Allosteric modulation of the 5-HT(3) receptor. (англ.) // Current Opinion In Pharmacology. — 2011. — February (vol. 11, no. 1). — P. 75—80. — doi:10.1016/j.coph.2011.01.010. — PMID 21342788.
- ↑ 1 2 3 Solt K., Stevens R. J., Davies P. A., Raines D. E. General anesthetic-induced channel gating enhancement of 5-hydroxytryptamine type 3 receptors depends on receptor subunit composition. (англ.) // The Journal Of Pharmacology And Experimental Therapeutics. — 2005. — November (vol. 315, no. 2). — P. 771—776. — doi:10.1124/jpet.105.090621. — PMID 16081679.