Апопто́з — регулируемый процесс программируемой клеточной гибели, в результате которого клетка распадается на отдельные апоптотические тельца, ограниченные плазматической мембраной. Фрагменты погибшей клетки обычно очень быстро фагоцитируются макрофагами либо соседними клетками, минуя развитие воспалительной реакции. Морфологически регистрируемый процесс апоптоза продолжается 1—3 часа. Одной из основных функций апоптоза является уничтожение дефектных клеток. В многоклеточных организмах апоптоз к тому же задействован в процессах дифференциации и морфогенеза, в поддержании клеточного гомеостаза, в обеспечении важных аспектов развития и функционирования иммунной системы. Апоптоз наблюдается у всех эукариотов, начиная от одноклеточных простейших и вплоть до высших организмов. В программируемой смерти прокариотов участвуют функциональные аналоги эукариотических белков апоптоза.
Протеинкина́зы — подкласс ферментов киназ (фосфотрансфераз). Протеинкиназы модифицируют другие белки путём фосфорилирования остатков аминокислот, имеющих гидроксильные группы или гетероциклической аминогруппы гистидина.
Транскрипцио́нный фактор NF-κB — универсальный фактор транскрипции, контролирующий экспрессию генов иммунного ответа, апоптоза и клеточного цикла. Нарушение регуляции NF-kB вызывает воспаление, аутоиммунные заболевания, а также развитие вирусных инфекций и рака. Семейство NF-kB состоит из 5 белков: NF-kB1, NF-kB2, RelA, RelB и c-Rel, образующих 15 комбинаций димеров. Все белки семейства объединяет наличие домена гомологии Rel, который обеспечивает образование белковых димеров, связывание NF-kB с ДНК и с цитозольным ингибиторным белком IkB. Фактор NF-kB проявляет активность только в димерной форме, причём наиболее распространённые формы — димеры субъединиц p50 или p52 с субъединицей p65.

Каспазы — протеолитические ферменты, относящихся к семейству цистеиновых протеаз, расщепляющих белки исключительно после аспартата. Каспазы играют важную роль в процессах апоптоза, некроза и воспалительных процессах.

B-клеточный рецептор, или B-клеточный рецептор антигена — мембранный рецептор В-клеток, специфично узнающий антиген. Фактически В-клеточный рецептор представляет собой мембранную форму антител (иммуноглобулинов), синтезируемых данным В-лимфоцитом, и имеет ту же субстратную специфичность, что и секретируемые антитела. Этот рецептор, как и антитела, может существовать в нескольких формах в зависимости от того, к какому классу принадлежат его тяжёлые цепи. С В-клеточного рецептора начинается цепь передачи сигнала внутрь клетки, которая в зависимости от условий может приводить к активации, пролиферации, дифференцировке или апоптозу В-лимфоцитов. Сигналы, поступающие от B-клеточного рецептора и его незрелой формы, оказываются критическими в созревании В-лимфоцитов и в формировании репертуара антител организма.
CD24, или передатчик сигнала CD24 — гликопротеин, который у человека кодируется геном CD24. CD24 состоит из короткой пептидной части и ГФИ-якоря, который удерживает его в клеточной мембране. Подвергается интенсивному тканеспецифичному гликозилированию и играет роль в адаптивном иммунном ответе, воспалении, аутоиммунных заболеваниях и злокачественном перерождении клеток.
Пре-B-клеточный рецептор — рецепторный белковый комплекс, который присутствует на плазматической мембране предшественников B-клеток в течение короткого времени и служит сигналом завершения продуктивной соматической рекомбинации генов тяжёлых цепей иммуноглобулинов. Хотя пре-В-клеточный рецептор синтезируется в клетке только в течение короткого промежутка времени, это событие, тем не менее, является важной контрольной точкой в развитии В-лимфоцита. Его присутствие на мембране показывает, что клетки способны синтезировать нормальные тяжёлые цепи иммуноглобулинов, и только такие клетки могут продолжать своё развитие, в противном случае они погибают в результате апоптоза. Синтез пре-B-клеточного рецептора знаменует переход от про-B-клеток к пре-B-клеткам.
BCL6, или белок 6 В-клеточной лимфомы, — фактор транскрипции, который у человека кодируется геном BCL6, протоонкоген. Гомологичные гены были обнаружены у мышей и крыс. BCL6 необходим для формирования герминативных центров лимфоидных фолликулов. Экспрессия BCL6 нарушена примерно в 40 % случаев диффузной B-крупноклеточной лимфомы и 5—10 % случаев фолликулярной лимфомы.
p66Shc – белок, участвующий в регуляции клеточного уровня активных форм кислорода (АФК), индукции апоптоза и продолжительности жизни. Характерен исключительно для позвоночных животных: он присутствует, например, у шпорцевой лягушки и млекопитающих, но отсутствует у дрозофилы и нематоды.
SMAD2 — белок, кодируемый у человека геном SMAD2. Он относится к семейству белков SMAD, члены которого подобны белковым продуктам гена Mad плодовой мушки Drosophila melanogaster и гена SMA нематоды C. elegans. Белки SMAD — преобразователи сигналов и транскрипционные модуляторы, которые опосредуют несколько сигнальных путей.

Daxx — белок, кодируемый у человека геном DAXX.

Сурвивин, также бакуловирусный ингибитор мотива апоптозных повторов 5, сокращённо BIRC5 — белок, кодируемый у человека геном BIRC5.
Некропто́з — программируемая некротическая гибель клетки, сопровождаемая активацией взаимодействующей с рецептором протеинкиназы-3. На молекулярном уровне при некроптозе происходит строго регулируемая сборка внутриклеточного комплекса, известного как некросома, запускаемая рецепторами смерти, поверхностными Toll-подобными рецепторами, а также механизмами, распознающими присутствие в цитоплазме вирусных РНК. Для некроптоза, индуцируемого фактором некроза опухоли (TNF), необходима дальнейшая активация RIPK1 (RIP1) и RIPK3. Блокирование этих киназ некростатинами, например, некростатином 1, ингибирующим RIPK1, делает некроптоз невозможным. В отличие от апоптоза, вызываемого активацией каспазы 8, некроптоз может протекать лишь при инактивации этого фермента. При некроптозе также происходит образование активных форм кислорода в митохондриях, однако, в отличие от апоптоза, не происходит фрагментация ДНК. Кроме того, в отличие от апоптоза, некроптоз сопровождается сильным иммунным ответом: погибающая клетка высвобождает молекулярные фрагменты, ассоциированные с повреждениями, которые активируют иммунитет. Некроптоз может запускаться в тех случаях, когда апоптоз по тем или иным причинам невозможен. В отличие от молекулярных путей апоптоза, изучаемых уже много лет, молекулярные основы некроптоза в настоящее время мало изучены.
Аноики́с — частный случай клеточной гибели путём апоптоза, которая наступает в ответ на неправильную адгезию клеток или её утрату. Правильная адгезия клетки ко внеклеточному матриксу необходима для того, чтобы понять, правильное ли место занимает клетка. Удаление клеток, потерявших связь со своим окружением, препятствует их связыванию с неправильным субстратом и дальнейшему росту. Таким образом, аноикис необходим для развития и поддержания гомеостаза тканей, а его неправильная регуляция имеет место при некоторых болезнях. Аноикис был описан у некоторых типов клеток, однако, судя по всему, клетки из различных тканей используют разные молекулярные пути, приводящие к аноикису.

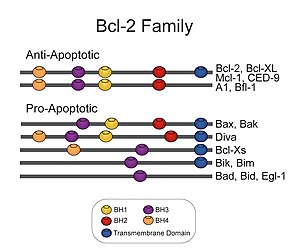
PUMA — про-апоптотический протеин из семейства Bcl-2.
Программи́руемая кле́точная ги́бель, или программи́руемая кле́точная смерть, или запрограмми́рованная ги́бель кле́ток — гибель клетки, которая происходит за счёт запрограммированных внутриклеточных процессов. Ко второму десятилетию XXI века насчитывается более десяти известных видов программируемой клеточной гибели. С 2005 года классификацией видов клеточной гибели занимается Комитет по номенклатуре видов клеточной гибели. Программируемая клеточная гибель описана для всех крупных групп эукариот: животных, растений, грибов, слизевиков и даже одноклеточных организмов. ПКГ выполняет множество функций как на уровне клетки, так и на уровне целого организма: у животных она играет важнейшую роль в развитии, с её помощью элиминируются повреждённые клетки, у растений она задействована в образовании тканей, состоящих из мёртвых клеток, таких как ксилема. Программируемая клеточная гибель известна не только у эукариот: несколько видов программируемой гибели было описано у бактерий. Все виды программируемой клеточной гибели можно подразделить на внешние, которые запускаются сигналами извне клетки, и внутренние, вызванные нарушениями в функционировании клеток.

Каспаза 3 — протеолитический фермент, одна из каспаз человека, взаимодействует с каспазой 8 и каспазой 9. Кодируется одноимённым геном — CASP3, который локализован на коротком плече (p-плече) 4-ой хромосомы. Ортологи CASP3 были идентифицированы у многих видов млекопитающих, для которых имеются полные данные генома. Уникальные ортологи также присутствуют у птиц, ящериц, лиссамфибий и телеостов. Длина полипептидной цепи белка составляет 277 аминокислотных остатков, а молекулярная масса — 31608 Да.

Каспаза 8 — протеолитический фермент, одна из каспаз, кодируется одноимённым геном CASP8, который у человека локализован на коротком плече (p-плече) 2-ой хромосомы. Является экзекуторной (эффекторной) каспазой. Скорее всего, действует на каспазу 3. Ортологи CASP8 были идентифицированы у многочисленных видов млекопитающих, для которых имеются полные данные генома. Эти уникальные ортологи также присутствуют у птиц.

Fas-ассоциированный белок с доменом смерти или белок, взаимодействующий с доменом смерти Fas-рецептора, также называемый MORT1 — адаптерный белок, кодируется одноимённым геном FADD, у человека он локализован на длинном плече (q-плече) 11-ой хромосомы.

Fas-рецептор, также известный как апоптозный антиген 1, кластер дифференцировки 95 (CD95) или член суперсемейства рецепторов фактора некроза опухолей 6 (TNFRSF6) — белок, который у людей кодируется геном FAS. Впервые Fas был идентифицирован с использованием моноклональных антител, полученных путём иммунизации мышей клеточной линией FS-7. Таким образом, название Fas происходит от FS-7-ассоциированного поверхностного антигена.