Генетическая инжене́рия — совокупность приёмов, методов и технологий получения рекомбинантных РНК и ДНК, выделения генов из организма (клеток), осуществления манипуляций с генами, введения их в другие организмы и выращивания искусственных организмов после удаления выбранных генов из ДНК. Генетическая инженерия не является наукой в широком смысле, но является инструментом биотехнологии, используя методы таких биологических наук, как молекулярная и клеточная биология, генетика, микробиология, вирусология.
Аммонификáция — процесс разложения азотсодержащих органических соединений в результате их ферментативного гидролиза под действием аммонифицирующих микроорганизмов с образованием токсичных для человека конечных продуктов — аммиака, сероводорода, а также первичных и вторичных аминов при неполной минерализации продуктов разложения:
- трупных ядов, например: путресцин и кадаверин;
- ароматических соединений, например: скатол, индол — образуются в результате дезаминирования и декарбоксилирования аминокислоты триптофана;
- серосодержащих аминокислот, например: цистеина, цистина и метионина, — приводит к выделению сероводорода, тиолов, диметилсульфоксида.
Химическая эволюция или пребиотическая эволюция — этап, предшествовавший появлению жизни, в ходе которого органические, пребиотические вещества возникли из неорганических молекул под влиянием внешних энергетических и селекционных факторов и в силу развертывания процессов самоорганизации, свойственных всем относительно сложным системам, которыми, бесспорно, являются все углеродосодержащие молекулы.
Хемосинтез — способ автотрофного питания, при котором источником энергии для синтеза органических веществ из CO2 служат реакции окисления неорганических соединений. Подобный вариант получения энергии используется только бактериями или археями. Это явление было открыто в 1887 году русским учёным С. Н. Виноградским. Микроорганизмов, способных к хемосинтезу, Виноградский называл аноргоксиданты. Название хемосинтез ввёл немецкий химик и ботаник Вильгельм Пфеффер в 1897 году.
Азотфикса́ция, или азотофиксация, — фиксация молекулярного атмосферного азота, диазотрофия. Процесс восстановления молекулы азота и включения её в состав своей биомассы прокариотными микроорганизмами. Важнейший источник азота в биологическом круговороте. В наземных экосистемах азотфиксаторы локализуются в основном в почве.
Денитрификация — сумма микробиологических процессов восстановления нитратов до нитритов и далее до газообразных оксидов и молекулярного азота. В результате их азот возвращается в атмосферу и становится недоступным большинству организмов. Осуществляется только прокариотами в анаэробных условиях и связана с получением ими энергии.

Анаэробы — организмы, получающие энергию при отсутствии доступа кислорода путём субстратного фосфорилирования, конечные продукты неполного окисления субстрата при этом могут быть окислены с получением большего количества энергии в виде АТФ.
Гетероцисты — дифференцированные клетки нитчатых цианобактерий, осуществляющие азотфиксацию. При недостатке соединений азота в среде они появляются регулярно вдоль трихомы из вегетативных клеток и акинет. Цианобактерии — фототрофы, осуществляющие оксигенный фотосинтез, однако кислород, атмосферный и выделяемый при фотосинтезе, ингибирует фермент нитрогеназу, необходимую для азотфиксации, поэтому у нитчатых цианобактерий в процессе эволюции возникли специализированные клетки для азотфиксации.
Анаэробное дыхание — это биохимический процесс окисления органических субстратов или молекулярного водорода с использованием в дыхательной ЭТЦ в качестве конечного акцептора электронов вместо O2 других окислителей неорганической или органической природы. Как и в случае аэробного дыхания, выделяющаяся в ходе реакции свободная энергия запасается в виде трансмембранного протонного потенциала, использующегося АТФ-синтазой для синтеза АТФ.

Анаммо́кс (сокр. от англ. anaerobic ammonium oxidation — анаэробное окисление аммония) — один из ключевых микробных процессов в круговороте азота. Бактерии, осуществляющие этот процесс, были открыты в 1999 году, и в своё время описание этого процесса стало большим сюрпризом для научного сообщества. Уравнение процесса:
- NH4+ + NO2− → N2 + 2H2O.

Круговорот азота — биогеохимический цикл азота. Большая его часть обусловлена действием живых существ. Важную роль в круговороте играют почвенные микроорганизмы, обеспечивающие азотистый обмен почвы — круговорот в почве азота, который присутствует там в виде простого вещества (газа — N2) и ионов: нитритов (NO2-), нитратов (NO3-) и аммония (NH4+). Концентрации этих ионов отражают состояние почвенных сообществ, поскольку на эти показатели влияет состояние биоты (растений, микрофлоры), состояние атмосферы, вымывание из почвы различных веществ. Они способны снижать концентрации азотсодержащих веществ, губительные для других живых организмов. Они могут переводить токсичный для живых существ аммиак в менее токсичные нитраты и в биологически инертный атмосферный азот. Таким образом, микрофлора почвы способствует поддержанию стабильности её химических показателей.
Альтернативная биохимия — ряд теорий и гипотез, рассматривающих возможность существования форм жизни частично или полностью отличающихся биохимически от возникших на Земле. Обсуждаемые в рамках гипотез отличия включают замену углерода в молекулах органических веществ на иные атомы, либо замену воды в качестве универсального растворителя другими жидкостями. Альтернативная биохимия нередко встречается в научно-фантастической литературе и спекулятивной биологии при описании различных форм внеземной жизни. Но также является предметом серьёзного научного интереса и исследований.
После́дний универса́льный о́бщий пре́док — последняя популяция организмов, от которой произошли все организмы, ныне живущие на Земле. Таким образом, LUCA является общим предком всей жизни на Земле. Последнего универсального общего предка не следует путать с первым живым организмом на Земле. Считается, что LUCA жил 3,48—4,28 миллиарда лет назад, или, возможно, даже 4,5 млрд лет назад. Ископаемых остатков LUCA не сохранилось, поэтому его можно изучать только путём сравнения геномов. С помощью этого метода в 2016 году был определён набор из 355 генов, точно имевшихся у LUCA.

Сульфатредуцирующие бактерии представляют собой группу бактерий, характеризующуюся способностью окислять сероводород и отлагать в своём теле крупинки серы. Признак этой группы, как видно из сказанного, чисто физиологический, морфологических же признаков эта группа не имеет. Известны учёным серные бактерии уже давно. Ф. Кон первый изучал их физиологию, и так как серные бактерии всегда были находимы в тех местах, где наблюдалось выделение сероводорода, то Кон пришёл к заключению, что образование сероводорода находится в зависимости от жизнедеятельности этих бактерий, разрушающих различные соединения, заключающие в себе серу, и выделяющих сероводород. В этом своём заключении он опирается на наблюдения Лотара Мейера, который заметил, что вода Линдекских минеральных источников с серными бактериями — «водорослями», как он думал — после четырёхмесячного стояния заключает гораздо больше сероводорода, чем без «водорослей», и что «водоросли» восстанавливают, по-видимому, сернокислые соли в сероводород. Наблюдения над минеральными источниками в Иоганисбаде показали, далее, Кону, что там, где нет серных бактерий (Beggiatoa), там нет и сероводорода, и наоборот; кроме того, исследуя материал, присланный ему с берегов Дании Вормигом из мест, где замечалось выделение сероводорода, Кон нашёл много спирилл и монад с отложением серы внутри их тела, что ещё более укрепило его во взгляде на способность многих микроорганизмов выделять сероводород. Гоппе-Зейлер первый усомнился в справедливости заключений Кона, а Виноградский блестяще доказал ошибочность взглядов Кона; взгляд Виноградского считается теперь общепризнанным. По его мнению, серные бактерии не имеют никакого отношения к образованию сероводорода и восстановлению сульфатов, отложение же серы внутри тела бактерий Виноградский принимает как следствие окисления находящегося в воде сероводорода бактериями. Шаблон:Биофото Для получения серных бактерий в культурах Виноградский разрезал на мелкие куски свежевыкопанное корневище водяного растения — лучше всего для этой цели ему служило корневище сусака — и клал их в высокий сосуд, в который наливал колодезной воды с прибавкой небольшого количества гипса. Уже по прошествии 5—6 дней вода сосуда начинала пахнуть сероводородом, выделение его понемногу увеличивалось, но серных бактерий заметно ещё не было. Только через 4 недели можно, наконец, заметить появление нитей Beggiatoa, а месяца через 2 стенки сосуда вблизи поверхности воды оказывались уже вполне ими покрыты. Если кусочки Butomus umbellatus перед тем, как бросить в воду, подержать некоторое время в кипящей воде, то хотя сероводород и образуется потом в сосуде с гипсом, но Beggiatoa не появляется. Таким образом Виноградский мог заключить, что не Beggiatoa образует сероводород, но газ этот появляется помимо серных бактерий. Сера отлагается в теле бактерий только тогда, когда бактерии развиваются в воде, заключающей сероводород, и исчезает уже по прошествии 24 часов, когда бактерии переносят в ключевую или кипячёную воду. В случае недостатка сероводорода в воде бактерии окисляют находящуюся в их клетках серу до серной кислоты, которая образует с углекислой известью окружающей воды гипс. Схематически процессы, происходящие при этом, можно представить в виде следующих формул:

 .
.
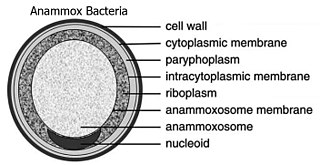
Candidatus Brocadia anammoxidans (лат.) — вид анаэробных хемолитоавтотрофных кокковидных бактерий рода Candidatus Brocadia, впервые выделен из сооружений по очистке сточных вод в Делфте, Нидерланды, также были обнаружены в Чёрном море.
Aigarchaeota — тип архей, родственный кренархеотам (Crenarchaeota) и пока не получивший признания во всех системах. К нему относят 1—2 вида: Candidatus Caldiarchaeum subterraneum и Candidatus Calditenuis aerorheumensis.

Зелёные серобактерии (лат. Chlorobiaceae) — семейство облигатно анаэробных (более строгих, чем пурпурные бактерии, в присутствии O2 не растут) фотолитоавтотрофных грамотрицательных бактерий, использующих сероводород (H2S), водород (H2) и элементарную серу (S0) в качестве доноров электронов. По происхождению они принадлежат к надтипу Bacteroidetes-Chlorobi, однако неоднородны и потому их классифицируют как отдельный тип.
Зелёные несе́рные бакте́рии, или зелёные ни́тчатые серобакте́рии, — филогенетически однородная группа факультативно анаэробных фотогетеротрофных бактерий, осуществляющих аноксигенный фотосинтез, использующих H2S, H2, сахара, аминокислоты и органические кислоты в качестве доноров электронов. В отличие от зелёных серобактерий, зелёные несерные бактерии при окислении сероводорода откладывают элементарную серу вне клеток, за что и получили своё название. Следует отметить, что зелёные несерные бактерии филогенетически довольно далеко отстоят от зелёных серобактерий. Изначально к этой группе относили весь класс Chloroflexia, но позднее такие представления были пересмотрены, когда выяснилось, что микроорганизмы порядка Herpetosiphonales не способны к фотосинтезу. Согласно современным представления, к этой группе относят представителей четырёх родов.
Ацетогены — прокариоты, которые выделяют ацетат (CH3COO−) в качестве конечного продукта анаэробного дыхания или гомоацетатного брожения. Однако обычно этот термин используется в более узком смысле: только в отношении бактерий и архей, которые одновременно осуществляют анаэробное дыхание и фиксацию углерода в восстановительном ацетил-КоА-пути (путь Вуда — Льюнгдаля). Этот процесс известен как ацетогенез. Ацетогенов следует отличать от уксуснокислых бактерий, которые выделяют ацетат, получающийся в результате неполного аэробного окисления этилового спирта до уксусной кислоты.
Сероводородное брожение — сложный биохимический процесс, состоящий в образовании сапрогенными микроорганизмами сероводорода и аммиака из сложных органических, а также неорганических, содержащих серу веществ и в дальнейших реакциях этих газов с различными солями, приходящими с ними в соприкосновение в окружающей среде — процесс существенный и грандиозный в мировом круговороте серы.

