Eusthenodon
| † Eusthenodon | |||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 Реконструкция | |||||||||||||||||||||||||||||||||||||||||
| Научная классификация | |||||||||||||||||||||||||||||||||||||||||
Домен: Царство: Подцарство: Без ранга: Без ранга: Тип: Подтип: Инфратип: Группа: Класс: Клада: Клада: Подкласс: † Osteolepidida Отряд: Семейство: Род: † Eusthenodon | |||||||||||||||||||||||||||||||||||||||||
| Международное научное название | |||||||||||||||||||||||||||||||||||||||||
| Eusthenodon Jarvik, 1952 | |||||||||||||||||||||||||||||||||||||||||
| Виды | |||||||||||||||||||||||||||||||||||||||||
| Геохронология 382,7—358,9 млн лет
◄ Наше время◄ Мел-палеогеновое вымирание ◄ Триасовое вымирание◄ Массовоепермское вымирание ◄ Девонское вымирание◄ Ордовикско-силурийское вымирание◄ Кембрийский взрыв | |||||||||||||||||||||||||||||||||||||||||
Eusthenodon (лат., от др.-греч. eusthenes — крепкий, и др.-греч. odon — зуб) — вымерший род тетраподоморф из семейства Tristichopteridae, обитавших в позднем девонском периоде, от франского до фаменского веков[3][4]. Космополитный род, ископаемые остатки известны из Восточной Гренландии, Австралии, средней полосы России, ЮАР, Пенсильвании и Бельгии[5][6]. При сравнении с родственными родами клады Tristichopteridae обнаруживается, что Eusthenodon был одной из крупнейших лопастепёрых рыб (около 2,5 м в длину) и одной из наиболее развитых в своём семействе наряду с Cabonnichthys и Mandageria[5][4].
Большие размеры, хищническая экология и эволюционно развитые признаки Eusthenodon, вероятно, способствовали его распространению и процветанию по всему миру. Eusthenodon считается лишь одним из многих космополитичных родов в ихтиофауне древнего красного песчаника верхнего девона[3][7][8]. В результате была выдвинута гипотеза, что диверсификация Eusthenodon и других морфологически сходных Tristichopteridae не сдерживалась биогеографическими барьерами, а ограничивалась только индивидуальной экологией и подвижностью животных[8].
Большинство остатков Eusthenodon, найденных в различных местах, состоят в основном из черепных элементов, в подавляющем большинстве случаев цельные скелеты отсутствуют[5][8][7]. Как следствие, основная часть доступной литературы, посвящённой Eusthenodon, в основном сосредоточена на проблемах, связанных с черепом, в то время как другие авторы делают выводы по признакам, типичным для Tristichopteridae[8]. Йохансон и Альберг (1997) в своей оценке нового материала саркоптеригий предполагают, что Eusthenodon, вероятно, обладал таким же трифуркатевидным или ромбовидным хвостовым плавником с осевой долей, слегка повёрнутой дорсально, что характерно для других Tristichopteridae (Йохансон именует их Eusthenopteridae), а также первым спинным плавником треугольной формы[5].
Открытие и наименование
В 1952 году шведский палеонтолог Эрик Ярвик[англ.] впервые описал первый вид Eusthenodon wangsjoi[9]. Образец был извлечён в 1936 году из богатых окаменелостями верхнедевонских наносов Восточной Гренландии — региона, который обрёл огромную привлекательность для исследователей древних позвоночных[англ.] после открытия обладавшей лапами Ichthyostega[10]. Родовое название Eusthenodon связано с хорошо различимыми бивнями на верзней и нижней челюстях[11].
Видовое название дано в честь первооткрывателя Густава Венгшё. Первоначально Ярвик писал его как wängsjöi. Однако МКЗН запрещает использование диакритических знаков в биномах. В 1962 году Воробьёва изменила видовое название на waengsjoei. В 2009 году Даниэль Сниттинг и Хеннинг Блом указали на то, что тут по ошибке присутствуют немецкие умлауты. В соответствии со статьёй 32.5.2.1 МКЗН правильное написание — wangsjoi[12].
Описание
Череп
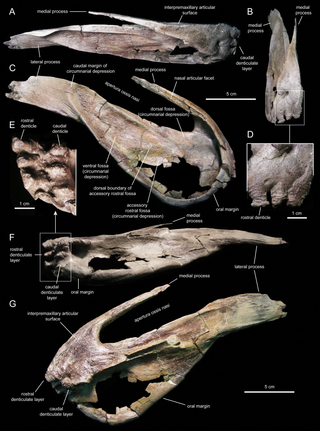
В своей первоначальной диагностике первых остатков Eusthenodon, опубликованной в 1952 году, Ярвик описывает особенности, присутствующие у Eusthenodon wangsjoi, в том числе те, которые являются значимыми признаками Tristichopteridae (названных Ярвиком Rhizodontida), а также а также признаки, уникальные для описываемого вида и диагностические признаки рода[9]. Заострённая голова Eusthenodon заметно крупнее, чем у родственных Osteolepiformes, теменные щитки короткие, из-за которых рыло широкое[13]. Лобно-этмоидальный щиток[англ.] черепной крыши заметно длиннее теменного щитка[11]. Соотношение длины лобно-этмоидального и теменного щитков использовалось палеонтологами для различения таксонов, и в некоторых случаях служит единственным отличительным признаком двух групп (как это видно при разделении клад Eusthenopteron и Tristichopterus)[7][5]. Среди Eusthenopteridae (Tristichopteridae) существует тенденция, показывающая всё более высокие значения этого соотношения у более производных родов, причём Eusthenodon обладает самым высоким значением с отношением лобно-этмоидального щита к теменному щиту — 2,30[7]. Дальнейшее увеличение длины рыла у многих видов четвероногих также может быть дополнительным доказательством тенденции возрастания длины лобно-этмоидальных щитков в последующих кладах, тесно связанных с Eusthenopteridae, включая поздних эопод[7]. Глазные отверстия заметно меньше по сравнению с более крупным лобно-решётчатым щитком[11][7]. Задняя надглазничная кость, расположенная за глазным отверстием, простирается вниз вдоль отверстия и контактирует со слёзной костью[9]. В отличие от других Osteolepiformes, тоже обладающих задней надглазничной костью, которая простирается вентрально вдоль глазничного отверстия, контакт со слёзной задней надглазничной костью является диагностическим признаком Eusthenodon и приводит к отделению яремной[англ.] и заглазничной костей от соприкасающимся глазнным отверстием[9].
Расположение и относительные размеры дополнительных окон у Eusthenodon, включая fenestra exonarina, шишковидное отверстие и теменное окно, являются дополнительными диагностическими признаками рода[11][7][4]. Треугольное теменное окно хорошо известно у Eusthenodon своим большим размером и характерным задним "хвостом"окна, приближающимся к заднему лобному краю или соприкасающимся с ним[11][7]. Напротив, теменное окно значительно меньше по размеру и расположено отчетливо сзади как от центра лобной, так и от заглазничной кости лобно-этмоидального щитка[11]. При взгляде на череп Eusthenodon в дорсальной проекции видно, что fenestra exonarina расположена высоко и сбоку в рыле[11].
Из трёх височных костей, составляющих теменной щиток, присутствующих у Osteolepiformes (межвисочная[англ.], надвисочная[англ.] и вневисочная кости), наличие вневисочной кости в «постспиракулярном» положении, определяемом как смещение кости из латерального положения по отношению к надвисочной в более задне-латеральное, является значимым и диагностическим признаком клады Tristichopteridae[7][9]. Вневисочная кость, присутствующая у Eusthenodon, примечательна своим полным постспиракулярным положением, благодаря которому отсутствует контакт между надвисочной и вневисочной костями — состояние, известное только у Eusthenodon[7]. Одна из теорий, объясняющих тенденцию, наблюдаемую в заднем смещении вневисочной кости у более развитых рыб, предполагает, что изменение пропорций головы способствовало более обтекаемой форме тела и повышало маневренность и скорость животных в водной среде[7].
Наружная щёчная пластина у Eusthenodon хорошо задокументирована, она в 3,5 раза длиннее теменного щитка, её длина в три раза превышает высоту[9]. Щёчная пластина и нижняя челюсть у Eusthenodon пропорционально значительно длиннее, чем у любых других Osteolepiformes. Нижняя челюсть Eusthenodon уменьшается в высоте на протяжении от заднего конца к переднему[9].
Зубной ряд
Как следует из названия, Eusthenodon имеет большие бивни, которые выступают из верхней и нижней челюстей[9]. В частности, вдоль средней линии рыла на предчелюстной кости появляются два больших и толстых зуба[5]. Судя по неполному материалу, собранному при обнаружении Eusthenodon, самые большие бивни были не менее 50 мм в длину[9]. Эти два зуба уплощены в передне-заднем направлении и имеют характерные острые режущие края[7][4]. В опубликованном Гаэлем Клементом в 2009 году исследовании скопления Tristichopteridae обнаружено, что увеличенные зубы преимущественно находились на одной линии с зубным рядом предчелюстной кости и не образовывали пары[8]. По этой причине увеличенные предчелюстные зубы описаны как «псевдоклыки», а не настоящие клыки, которые, как считалось ранее, присутствовали у Eusthenodon[8]. Анализ горизонтального поперечного сечения первого клыка выявляет простой и неравномерно сложенный ортодентин[5]. Он обнаруживается в пульповой полости[7][5]. Наличие увеличенных псевдоклыков на предчелюстной кости у Eusthenodon подтвердило его филогенетическое положение в кладе Tristichopteridae, поскольку аналогичные зубные ряды[англ.] обнаружены у других развитых представителей этой группы[8]. Количество маленьких заострённых зубов вдоль зубного ряда ещё больше поддерживает тенденции зубного ряда с течением времени, поскольку у более производных родов обнаруживается большее количество зубов по сравнению с более примитивными видами наподобие Eusthenopteron[7].
Несмотря на наличие наборов предчелюстных псевдоклыков, у Eusthenodon и других крупных, филогенетически развитых Tristichopteridae присутствуют сложные передние зубы и характерные увеличенные клыки на нижней челюсти[5][8]. Слабовогнутое зубчатое поле парасфеноидной кости[англ.] приподнято у примитивных Tristichopteridae, в то время как у Eusthenodon оно заметно пониженное. Кроме того, наличие характерной лопастевидной вертикальной пластинки, присутствующей на переднем короноиде[англ.], существует у большинства других Tristichopteridae, но отсутствует у производных родов, таких как Eusthenodon[8]. У Tristichopteridae передний и средний короноиды оснащены как минимум одной парой клыков, в то время как у Eusthenodon задний короноид с двумя парами клыков[5][8]. Кроме того, известно, что расположенные с краю короноидные зубы присутствуют практически у всех других Tristichopteridae (отсутствуют они только у Cabonnicthys), однако у Eusthenodon и близкородственной Mandageria нет выдающихся краевых короноидных зубов вдоль передней части челюсти[7]. Редукция этих зубов указывает на филогенетическую близость Eusthenodon, Mandageria и Cabonnichthys и является производной характеристикой поздних Tristichopteridae[7]. У Eusthenodon присутствует небольшая парасимфизарная пластинка, прикреплённая к передней части мезиальной пластинки у пластинчатой кости[англ.][8][9]. Форма и размер парасимфизарной пластинки у Eusthenodon характерны для всех Tristichopteridae и являются диагностической характеристикой семейства[8][9].
Чешуя
В соответствии с особенностями, описанными Бергом (1955) как значимые диагностические признаки Tristichopteridae, Eusthenodon обладает пропорционально большими, отчётливо округлыми чешуйками без космина[англ.], с сетчатым рисунком гребней с редкими одиночными бугорками[14][9][15]. Вдобавок у каждой из этих круглых косминных чешуек имеется проксимальный центральный выступ для прикрепления, что также является диагностическим признаком Tristichopteridae[7][9]. В отличие от большинства других Tristichopteridae на чешуях Eusthenodon присутствуют гребни, образующие отчётливые сети, в то время как чешуйки Eusthenopteron как правило с более короткими гребнями в неполностью сросшихся бугорках[9][7]. Площадь перекрытия чешуек у Eusthenodon также больше, чем у чешуи Eusthenopteron[9].
Классификация
Eusthenodon принадлежит к семейству Tristichopteridae отряда Osteolepiformes в большом классе лопастепёрые (Sarcopterygii)[7]. Лопастепёрые — основная клада, которая отделилась от лучепёрых (Actinopterygii) с эволюцией лопастных плавников.
Филогения Tristichopteridae была описана Гаэлем Клементом, Дэниелом Сниттингом и П. Е. Альбергом (2008) после выявления минимально инклюзивной клады[8]:
| Osteolepiformes |
| |||||||||||||||||||||||||||||||||||||||||||||||||||
Примечания
- ↑ Downs, J. P.; Barbosa, J.; Daeschler, E. B. (2021). "A new species of Eusthenodon (Sarcopterygii, Tristichopteridae) from the Upper Devonian (Famennian) of Pennsylvania, U.S.A., and a review of Eusthenodon taxonomy". Journal of Vertebrate Paleontology (англ.). 41 (3): e1976197. Bibcode:2021JVPal..41E6197D. doi:10.1080/02724634.2021.1976197. S2CID 240453731.
- ↑ Downs, J. P.; Osatchuck, M. M.; Goodchild, O. A.; Daeschler, E. B. (2023). "Second species of Eusthenodon (Tristichopteridae, Sarcopterygii) from the Upper Devonian (Famennian) Catskill Formation of Pennsylvania, U.S.A., and a review of global Eusthenodon occurrence". Journal of Vertebrate Paleontology (англ.). doi:10.1080/02724634.2023.2201627.
- ↑ 1 2 Blom, Henning; Clack, Jennifer; Ahlberg, Per. (2007). "Devonian vertebrates from East Greenland: A review of faunal composition and distribution". Geodiversitas (англ.). 29: 119—132 — ResearchGate.
- ↑ 1 2 3 4 Clement, Gael (2002). "Large Tristichopteridae (Sarcopterygii, Tetrapodomorpha) from the Late Famennian Evieux Formation of Belgium". Palaeontology (англ.). 45 (3): 577—593. Bibcode:2002Palgy..45..577C. doi:10.1111/1475-4983.00250. ISSN 0031-0239.
- ↑ 1 2 3 4 5 6 7 8 9 10 Ahlberg, Per E.; Johanson, Zerina (1997-12-15). "Second tristichopterid (Sarcopterygii, Osteolepiformes) from the Upper Devonian of Canowindra, New South Wales, Australia, and phylogeny of the Tristichopteridae". Journal of Vertebrate Paleontology (англ.). 17 (4): 653—673. Bibcode:1997JVPal..17..653A. doi:10.1080/02724634.1997.10011015. ISSN 0272-4634.
- ↑ Lebedev, O A; Zakharenko, G V; Silantiev, V V; Evdokimova, I O (2018). "New finds of fishes in the lower uppermost Famennian (Upper Devonian) of Central Russia and habitats of the Khovanshchinian vertebrate assemblages". Estonian Journal of Earth Sciences (англ.). 67 (1): 59. doi:10.3176/earth.2018.04. ISSN 1736-4728.
- ↑ 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 Borgen, Ulf J.; Nakrem, Hans A. (2016-09-30). "Morphology, phylogeny and taxonomy of osteolepiform fish". Fossils and Strata. Fossils and Strata Series (англ.). 61: 1—481. doi:10.1002/9781119286448.ch1. ISBN 9781119286431. ISSN 2637-6032.
- ↑ 1 2 3 4 5 6 7 8 9 10 11 12 13 CLEMENT, GAËL; SNITTING, DANIEL; AHLBERG, PER ERIK (2009). "A New Tristichopterid (Sarcopterygii, Tetrapodomorpha) from the Upper Famennian Evieux Formation (Upper Devonian) of Belgium" (PDF). Palaeontology (англ.). 52 (4): 823—836. Bibcode:2009Palgy..52..823C. doi:10.1111/j.1475-4983.2009.00876.x. ISSN 0031-0239. Архивировано (PDF) 20 июля 2018.
- ↑ 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 Jarvik, Erik. On the fish-like tail in the ichthyostegid stegocephalians : with descriptions of a new stegocephalian and new crossopterygian from the Upper Devonian of East Greenland. : [англ.]. — C. A. Reitzel, 1952. — Vol. 114. — P. 54–68.
- ↑ Jarvik, Erik. On the fish-like tail in the ichthyostegid stegocephalians : with descriptions of a new stegocephalian and new crossopterygian from the Upper Devonian of East Greenland. : [англ.]. — C. A. Reitzel, 1952. — Vol. 114. — P. 6.
- ↑ 1 2 3 4 5 6 7 Jarvik, Erik. On the fish-like tail in the ichthyostegid stegocephalians : with descriptions of a new stegocephalian and new crossopterygian from the Upper Devonian of East Greenland. : [англ.]. — C. A. Reitzel, 1952. — Vol. 114. — P. 54.
- ↑ Daniel Snitting & Henning Blom, 2009, «Correcting Taxon Names Containing Diacritics—Examples from Paleozoic Vertebrates», Journal of Vertebrate Paleontology, 29(1): 269—270
- ↑ Jarvik, Erik. On the fish-like tail in the ichthyostegid stegocephalians : with descriptions of a new stegocephalian and new crossopterygian from the Upper Devonian of East Greenland. : [англ.]. — C. A. Reitzel, 1952. — Vol. 114. — P. 55.
- ↑ Johanson, Z.; Ritchie, A. (2000-01-01). "Rhipidistians (Sarcopterygii) from the Hunter Siltstone (Late Famennian) near Grenfell, NSW, Australia". Fossil Record (англ.). 3 (1): 111—136. doi:10.5194/fr-3-111-2000. ISSN 2193-0074.
- ↑ Berg, L. S. Translation of pp. 161-288 of System der rezenten und fossilen Fischartigen und Fische by Berg 1955 : [англ.]. — 1958.
Ссылки
- Eusthenodon (англ.) информация на сайте Paleobiology Database.