
Хромосо́мы — нуклеопротеидные структуры в ядре эукариотической клетки, в которых сосредоточена бо́льшая часть наследственной информации и которые предназначены для её хранения, реализации и передачи. Хромосомы чётко различимы в световом микроскопе только в период митотического или мейотического деления клетки. Набор всех хромосом клетки, называемый кариотипом, является видоспецифичным признаком, для которого характерен относительно низкий уровень индивидуальной изменчивости.

Плазми́ды — небольшие молекулы ДНК, физически обособленные от хромосом и способные к автономной репликации. Главным образом плазмиды встречаются у бактерий, а также у некоторых архей и эукариот. Чаще всего плазмиды представляют собой двухцепочечные кольцевые молекулы. Несмотря на способность к размножению, плазмиды, как и вирусы, не рассматриваются в качестве живых организмов.

Мейо́з, или редукционное деление — деление ядра эукариотической клетки с уменьшением числа хромосом в два раза. Происходит в два этапа. В результате мейоза образуются гаметы, споры и другие зародышевые клетки.
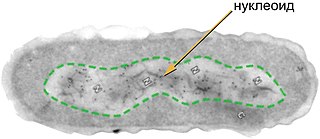
Нуклео́ид — неправильной формы зона в цитоплазме прокариотической клетки, в которой находится геномная ДНК и ассоциированные с ней белки. На долю ДНК приходится около 60 % массы нуклеоида; помимо ДНК, нуклеоид содержит РНК и белки. Белки нуклеоида, которые обеспечивают пространственную организацию геномной ДНК, называют нуклеоидными белками или нуклеоид-ассоциированными белками; они не имеют ничего общего с гистонами, упаковывающими ДНК у эукариот. В отличие от гистонов, ДНК-связывающие белки нуклеоида не формируют нуклеосомы и обеспечивают компактизацию геномной ДНК другим способом. Несмотря на аморфную форму, отдельные гены располагаются в нём упорядоченно.

Рекомбинация — перераспределение генетического материала путём разрыва и соединения разных молекул, приводящее к появлению новых комбинаций генов или других нуклеотидных последовательностей. В широком смысле слова включает в себя не только рекомбинацию между молекулами ДНК, но и перекомбинацию (сортировку) генетического материала на уровне целых хромосом или ядер, а также обмен плазмидами между клетками.

Франсуа́ Жако́б — французский микробиолог и генетик, нобелевский лауреат (1965). Член Французской академии (1996), иностранный член Национальной академии наук США (1969) и Лондонского королевского общества (1973).
Трансду́кция — процесс переноса ДНК между клетками при помощи вирусов. Примером трансдукции является перенос бактериальной ДНК из одной клетки в другую бактериофагом. Общая трансдукция используется в генетике бактерий для картирования генома. К трансдукции способны как умеренные фаги, так и вирулентные, последние, однако, уничтожают популяцию бактерий, поэтому трансдукция с их помощью не имеет большого значения ни в природе, ни при проведении исследований.
Конъюга́ция — однонаправленный перенос части генетического материала при непосредственном контакте двух бактериальных клеток. Открыт в 1946 году Джошуа Ледербергом и Эдвардом Татумом. Явление конъюгации было открыто и хорошо изучено у кишечной палочки, но в дальнейшем конъюгация была описана у множества как грамположительных, так и грамотрицательных бактерий. Посредством конъюгации бактерии обмениваются генетическим материалом, поддерживая своё генетическое разнообразие.
Эписомы — генетические элементы бактерий, способные существовать как в интегрированном с бактериальными хромосомами состоянии, так и в виде автономных плазмид.
Комплементарная ДНК — это ДНК, синтезированная на матрице зрелой мРНК в реакции, катализируемой обратной транскриптазой.

Профа́г — геном фага, интегрированный в хромосомную ДНК бактериальных клеток. Умеренные фаги интегрируются в геном клетки-хозяина или существуют в виде плазмид. Это латентная форма взаимодействия фага и бактериальной клетки, при которой не происходит лизис бактерий. При наличии повреждения клетки-хозяина начинается профаговая индукция, что ведет к старту литического цикла.
Реплико́н — молекула или участок ДНК или РНК, реплицирующийся из одной точки начала репликации.
Искусственная бактериальная хромосома — векторная система на основе F-плазмиды E. coli, участков cos фага лямбда и loxP фага Р1, используемая для клонирования длинных последовательностей ДНК. F-плазмида кодирует гены, регулирующие репликацию и контролирующие копийность. По участку loxP плазмидная ДНК может быть расщеплена белком Cre фага Р1, по cos-участку — соответствующим ферментом фага лямбда. Схожая векторная система под названием PAC была сделана на основе бактериальной P1-плазмиды из ДНК фага P1.
Фазмиды — молекулярные векторы, являющиеся искусственными гибридами между фагом и плазмидой. Фазмиды после встройки чужеродной ДНК могут в одних условиях развиваться как фаги, а в других как плазмиды.
А́нти-CRISPR — система белков, благодаря которой бактериофаги противостоят разрушительному действию систем CRISPR/Cas. Системы анти-CRISPR описаны у многих бактериофагов. Белки этих систем в большинстве случаев мешают процессу узнавания мишени и работе белков Cas. Системы анти-CRISPR могут иметь биотехнологическое значение, поскольку могут применяться для тонкой регуляции редактирования генома с помощью технологии CRISPR/Cas9.
R-плазми́да, или R-фа́ктор — плазмида резистентности, которые обеспечивает бактерии устойчивостью к антибиотикам. R-плазмиды были описаны примерно тогда же, когда антибиотики нашли широкое применение. Резистентность к новому лекарственному препарату может появиться в течение пяти лет после начала его применения. Появились и штаммы бактерий, обладающие резистентностью к нескольким антибиотикам одновременно; наиболее часто такие штаммы выявляются в больницах. Распространение множественной лекарственной устойчивости обусловлено широким применением антибиотиков в животноводстве и здравоохранении.

Ti-плазми́да — плазмида почвенной бактерии Agrobacterium tumefaciens, с помощью которой она вызывает опухоли у растений. Участок Ti-плазмиды, известный как T-ДНК, может встраиваться в геном растений и содержит гены биосинтеза фитогормонов и опинов, которые запускают образование опухоли.
Фаг P1 — умеренный бактериофаг, поражающий кишечную палочку и некоторые другие бактерии. При прохождении лизогенного цикла геном фага существует в виде плазмиды в бактерии, в отличие от других фагов, которые интегрируются в ДНК хозяина. P1 имеет икосаэдрическую головку, содержащую ДНК, прикреплённую к сократительному хвосту с шестью хвостовыми волокнами. Фаг P1 привлёк интерес исследователей, потому что его можно использовать для переноса ДНК из одной бактериальной клетки в другую в процессе, известном как трансдукция. При репликации во время своего литического цикла он захватывает фрагменты хромосомы хозяина. Если полученные вирусные частицы используются для заражения другого хозяина, захваченные фрагменты ДНК могут быть интегрированы в геном нового хозяина. Этот метод генной инженерии in vivo широко использовался в течение многих лет и используется до сих пор, хотя и в меньшей степени. P1 также можно использовать для создания производного от P1 вектора клонирования искусственной хромосомы, который может нести относительно большие фрагменты ДНК. P1 кодирует сайт-специфическую рекомбиназу Cre, которая широко используется для проведения клеточно-специфичной или специфичной по времени рекомбинации ДНК путем фланкирования ДНК-мишени сайтами loxP.
Фосмиды — это разновидность плазмид. Фосмиды, в отличие от космид, основаны на бактериальной F-плазмиде. Вектор клонирования ограничен, так как хозяин может содержать только одну молекулу фосмид. Фосмиды могут содержать вставки ДНК размером до 40 т.п.н.; часто источником вставки является случайная геномная ДНК. Фосмидную библиотеку готовят путем выделения геномной ДНК из организма-мишени и её клонирования в фосмидный вектор. Смесь для лигирования затем упаковывается в фаговые частицы, и ДНК трансфицируется в бактериального хозяина. Бактериальные клоны размножают библиотеку фосмида. Низкое число копий обеспечивает более высокую стабильность, чем векторы с относительно большим числом копий, включая космиды. Фосмиды могут быть полезны для создания стабильных библиотек из сложных геномов. Фосмиды обладают высокой структурной стабильностью, и было обнаружено, что они эффективно поддерживают ДНК человека даже после 100 поколений роста бактерий. Фосмидные клоны использовались для оценки точности общедоступной последовательности генома человека.