
Рибонуклеи́новая кислота́ (РНК) — одна из трёх основных макромолекул, которые содержатся в клетках всех живых организмов и играют важную роль в кодировании, прочтении, регуляции и экспрессии генов.

Ген — в классической генетике — наследственный фактор, который несёт информацию об определённом признаке или функции организма, и который является структурной и функциональной единицей наследственности. В таком качестве термин «ген» был введён в 1909 году датским ботаником, физиологом растений и генетиком Вильгельмом Йоханнсеном.
IRES — регуляторный участок мРНК эукариот и их вирусов, который обеспечивает кэп-независимую, или внутреннюю инициацию трансляции. При таком механизме инициации рибосома связывается с мРНК непосредственно в районе IRES, которые чаще всего располагаются в 5'-нетранслируемой области (5'-НТО) недалеко от сайта инициации трансляции, минуя стадии узнавания кэпа и сканирования.

Ма́тричная рибонуклеи́новая кислота́ — РНК, содержащая информацию о первичной структуре белков. мРНК синтезируется на основе ДНК в ходе транскрипции, после чего, в свою очередь, используется в ходе трансляции как матрица для синтеза белков. Тем самым мРНК играет важную роль в «проявлении» (экспрессии) генов.

Ми́кроРНК — малые некодирующие молекулы РНК длиной 18—25 нуклеотидов, обнаруженные у растений, животных и некоторых вирусов, принимающие участие в транскрипционной и посттранскрипционной регуляции экспрессии генов путём РНК-интерференции. Помимо внутриклеточной обнаружена внеклеточная (циркулирующая) микроРНК.
Некодирующая ДНК или Мусорная ДНК — части геномной ДНК организмов, которые не кодируют последовательности белков. Некоторые некодирующие ДНК переводятся в функциональные некодирующие РНК-молекулы. Другие функции некодирующей ДНК включают регуляцию последовательностей кодирующих белки, центромер и теломер. Термин «мусорная ДНК» стал популярным в 1960-х. В соответствии с T. Ryan Gregory, геномным биологом, первое явное обсуждение природы «мусорной» ДНК было сделано David Comings в 1972 году и он применил этот термин ко всем некодирующим ДНК. Термин был формализован Сусуму Оно в 1972 году, который заметил, что генетический груз нейтральных мутаций находится на верхнем пределе значений для функционирующих локусов, которые могли быть ожидаемыми исходя из типичной частоты мутаций. Сусуму предсказал, что геномы млекопитающих не могут содержать более чем 30 000 локусов из-за давления естественного отбора, так как «стоимость» мутационной нагрузки вызвала бы неизбежное снижение приспособленности, и в конечном счете вымирание. Этот прогноз остается верным, геном человека содержит приблизительно 20 000 генов. Другим подтверждением теории Оно служит наблюдение, что даже близкородственные виды могут иметь очень разные по размеру геномы, которое окрестили C-парадокс в 1971 году.

Альтернати́вный спла́йсинг — вариант сплайсинга матричных РНК (мРНК), при котором в ходе экспрессии гена на основе одного и того же первичного транскрипта (пре-мРНК) происходит образование нескольких зрелых мРНК. Структурные и функциональные различия образовавшихся транскриптов могут быть вызваны как выборочным включением в зрелую мРНК экзонов первичного транскрипта, так и сохранением в ней частей интронов. Наиболее распространённая разновидность альтернативного сплайсинга предусматривает пропуск экзона: отдельные экзоны транскрипта при определённых условиях могут быть как включены в зрелую мРНК, так и пропущены.

Антисмысловы́е РНК — одноцепочечные РНК, которые комплементарны мРНК, транскрибируемой в клетке, или гену-мишени. Механизмы действия антисмысловых РНК весьма разнообразны, они могут как подавлять, так и активировать экспрессию гена-мишени. Природные антисмысловые РНК есть и у прокариот, и у эукариот; они относятся к длинным некодирующим РНК как РНК длиной более 200 нуклеотидов. Синтетические антисмысловые РНК нашли широкое применение у исследователей в качестве инструмента для нокдауна генов. Антисмысловые РНК также находят медицинское применение.
Цис-регуляторные элементы — участки ДНК или РНК, регулирующие экспрессию генов, находящихся на той же молекуле.

SECIS-элеме́нт — участок РНК длиной около 60 нуклеотидов, формирующий шпилькообразную структуру. Этот структурный мотив заставляет стоп-кодон UGA кодировать селеноцистеин. Поэтому элемент SECIS является неотъемлемым элементом мРНК, кодирующих селенопротеины.

Шпи́лька — в молекулярной биологии элемент вторичной структуры РНК, а также одноцепочечной ДНК. Шпилька образуется в том случае, когда две последовательности одной и той же цепи комплементарны друг другу и соединяются друг с другом, перегибаясь одна к другой и образуя на конце неспаренный участок — петлю. Такие комплементарные последовательности нередко представляют собой палиндромные последовательности.
Нетрансли́руемые о́бласти — особые участки мРНК, не выступающие в качестве матрицы для синтеза белка и прилегающие с обеих сторон к транслируемой области. Таких области две: 5'-нетранслируемая область, или 5'-НТО и 3'-нетранслируемая область, или 3'-НТО, располагающиеся на 5'- и 3'-конце мРНК соответственно. Такое же название имеют участки ДНК, соответствующие 5'-НТО и 3'-НТО транскрипта.
5′-Нетранслируемая область, или лидерная последовательность — некодирующий участок мРНК, располагающийся сразу после кэпа, но перед кодирующей областью. Такое же название имеет участок ДНК, соответствующий 5′-НТО транскрипта. В 5′-НТО располагаются различные элементы, принимающие участие в регуляции эффективности трансляции.
AU-бога́тые элеме́нты — регуляторные мотивы, располагающиеся в 3'-UTR мРНК некоторых генов и играющие ключевую роль в стабилизации транскриптов этих генов. Эти элементы имеют размер от 50 до 150 нуклеотидов и обычно содержат многочисленные копии пентануклеотида AUUUA.

Железозави́симый элеме́нт (англ. Iron response element, IRE) — особый регуляторный элемент, содержащийся в 5′-нетранслируемой области мРНК белков, как правило, участвующих в метаболизме железа. Наиболее известным белком, мРНК которого содержит IRE, является ферритин, связывающий Fe3+. Регуляция, осуществляемая IRE, зависит от концентрации железа в клетке. IRE встречаются у самых разнообразных эукариот, но мало представлены среди растений.
3′-Нетранслируемая область — некодирующий участок мРНК, располагающийся на её 3′-конце после кодирующей области. Такое же название имеет участок ДНК, соответствующий 3′-UTR транскрипта. 3′-UTR может принимать участие в регуляции эффективности трансляции, стабильности мРНК, содержать сигналы полиаденилирования и сайты связывания микроРНК, а также выполнять ряд других регуляторных функций.

Рибопереключа́тель — элемент, как правило, 5'-нетранслируемой области (5'-НТО) мРНК. Он осуществляет цис-регуляцию мРНК, на которой находится, путём связывания с лигандами — разнообразными малыми молекулами, например, кобаламидом, тиаминпирофосфатом, лизином, глицином, флавинмононуклеотидом, гуанином, аденином и другими. Типичный рибопереключатель включает два основных домена: аптамерный домен, осуществляющий распознавание лиганда и связывание с ним, и платформу экспрессии, которая взаимодействует с белками транскрипции или трансляции. Аптамерный домен и платформа экспрессии перекрываются в области так называемой переключающей последовательности, которая и отвечает за сворачивание РНК в две взаимноисключающие вторичные структуры, за счёт чего и осуществляется регуляция.

Внутренняя терминация, также ρ-независимая терминация, ро-независимая терминация , — механизм остановки транскрипции гена у прокариот. Суть этого механизма такова: мРНК содержит гуанин-цитозин-обогащённую последовательность, которая может образовывать структуры типа шпилька в 7—20 пар оснований в длину. Гуанин и цитозин образуют друг с другом три водородные связи и поэтому связаны довольно прочно. Сразу после шпильки располагается участок, обогащённый урацилом. Связи между урацилом и аденином очень слабы. Белок, связанный с РНК-полимеразой (nusA), настолько прочно связывается со шпилькой, что это вызывает временную остановку полимеразы и прекращение транскрипции. В этот момент полимераза располагается на полиурациловом участке последовательности. Слабые аденин-урациловые связи снижают энергию дестабилизации дуплекса РНК-ДНК, что позволяет им ослабить напряжение полинуклеотидной цепи и диссоциировать от РНК-полимеразы.
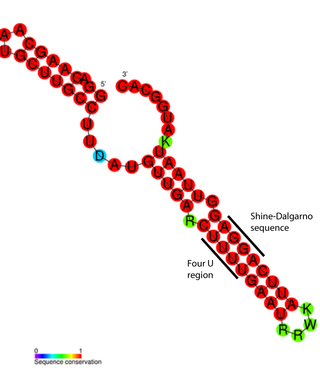
РНК-термо́метр, или РНК-термосе́нсор — температурочувствительная некодирующая РНК, которая принимает участие в регуляции экспрессии генов. РНК-термометры, как правило, регулируют гены, которые необходимы для ответа на тепловой или холодовой шок, однако показано их участие в регуляции длительного голодания и патогенности.
Старт-кодон или инициаторный кодон — первый кодон матричной РНК, c которого начинается трансляция белка в рибосоме. У эукариот и архей старт-кодон всегда кодирует метионин, а у прокариот— модифицированный метионин (N-формилметионин). В большинстве случаев роль инициаторного кодона играет триплет AUG. Старт-кодону предшествует 5′-нетранслируемая область (5'-UTR). В 5'-UTR бактерий локализована последовательность Шайна — Дальгарно (AGGAGG), которая служит для связывания рибосомы и отделёна спейсером от старт-кодона.