
Белки́ — высокомолекулярные органические вещества, состоящие из альфа-аминокислот, соединённых в цепочку пептидной связью. В живых организмах аминокислотный состав белков определяется генетическим кодом, при синтезе в большинстве случаев используется 20 стандартных аминокислот. Множество их комбинаций определяет большое разнообразие свойств молекул белков. Кроме того, аминокислотные остатки в составе белка часто подвергаются посттрансляционным модификациям, которые могут возникать и до того, как белок начинает выполнять свою функцию, и во время его «работы» в клетке. Часто в живых организмах несколько молекул разных белков образуют сложные комплексы, например фотосинтетический комплекс и другие комплексы.

Цитохром — небольшой гем-содержащий белок, относится к классу цитохромов, содержит в структуре гем типа c. Выполняет в клетке две функции. С одной стороны, он является одноэлектронным переносчиком, свободно связанным с внутренней мембраной митохондрий, и необходимым компонентом дыхательной цепи. Он способен окисляться и восстанавливаться, но не связывает при этом кислород. С другой стороны, при определённых условиях он может отсоединяться от мембраны, переходить в раствор в межмембранном пространстве и активировать апоптоз. Такая двойственность связана со специфичными свойствами молекулы цитохрома c.
Протеинкина́зы — подкласс ферментов киназ (фосфотрансфераз). Протеинкиназы модифицируют другие белки путём фосфорилирования остатков аминокислот, имеющих гидроксильные группы или гетероциклической аминогруппы гистидина.

НАДФH-оксидаза, или NADPH-оксидаза (NOX) — клеточный мембрано-связанный мультимолекулярный ферментный комплекс, локализующийся на плазматической мембране и в некоторых органеллах. Особенно обогащены этим ферментом фагоцитарные клетки, такие как макрофаги. Эти оксидазы участвуют в клеточной противомикробной защитной системе, а также в клеточной пролиферации, дифференцировке и регуляции экспрессии генов. Существует целая группа НАДФH-оксидаз, которые различаются по составу субъединиц, клеточной специфичности, регуляции и другим параметрам.
NADPH-оксидаза 4 (NOX4) — тип NADPH-оксидазы, клеточной мембранной оксидоредуктазы, образующей супероксидный радикал при переносе электрона с NADPH на кислород. В отличие от других, форм, требующих для активации образования мультибелкового комплекса с несколькими цитозольными белками, является конститутивно активной.

Цитохром b-245, альфа-полипептид, или p22-phox — трансмембранный белок, субъединица цитохрома b, состоящего из лёгкой цепи (альфа) и тяжёлой цепи (бета). Ген CYBA кодирует лёгкую альфа-субъединицу, необходимый компонент практически всех NADPH-оксидаз. Образует гетеродимер с тяжёлой цепью цитохрома, основным каталитическим компонентом NADPH-оксидаз, требующих, однако, для активации взаимодействия с несколькими регуляторными белками. Играет роль в продукции супероксидов и фагоцитозе.

NADPH-оксидаза 2, или gp91phox — основной фагоцитарный тип NADPH-оксидазы, клеточной мембранной оксидоредуктазы. Продуцирует супероксидный радикал, необходимый нейтрофилам и макрофагам для уничтожения микроорганизмов.

Цитозольный фактор нейтрофилов 1, или p47-phox — цитозольный регуляторный белок молекулярной массой 47-кДа комплекса NADPH-оксидазы 2. Цитохром b, состоящий из лёгкой цепи (альфа) и тяжёлой цепи (бета) требует связывание этого белка для своей каталитической активации.
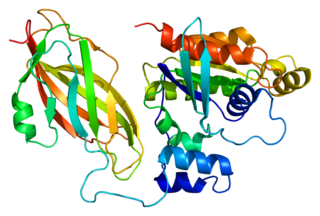
PECAM-1, или CD31, — гликопротеин, мембранный белок из суперсемейства иммуноглобулинов, относится к классу молекул клеточной адгезии. Один из основных белков межклеточных контактов эндотелиальных клеток: одна клетка может содержать до 1 млн копий белка. Часто используется как маркёр эндотелия при иммуногистологическом анализе ткани. Обнаружен также в ме́ньших количествах на циркулирующих тромбоцитах, моноцитах, нейтрофилах и на некоторых типах Т-клеток. Он также относится к подгруппе белков с ингибиторным ITIM участком. Всего известно 30 ITIM-содержащих белков.
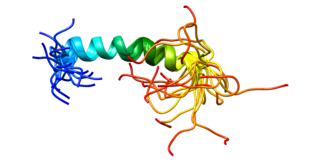
Rac2 — цитозольный белок, Rho ГТФаза из суперсемейства ГТФаз, относится к «малым» G-белкам. Находится в двух состояниях: активном ГТФ-связанном и неактивном ГДФ-связанном состоянии. В своей активной форме связывается в клетке с целым рядом эффекторных белков и приводит к регулировке многих клеточных процессов, таких как секреция, фагоцитоз апоптозных клеток, поляризация эпителиальных клеток и индуцированное факторами роста образование мембранных складок и выростов. Участвует в активации NADPH-оксидазы NOX2.

Интегрин бета-2 (CD18) — мембранный белок, гликопротеин из надсемейства интегринов, продукт гена ITGB2.

Интегрин альфа-5 — мембранный белок, гликопротеин из надсемейства интегринов, продукт гена ITGA5, альфа-субъединица интегрина α5β1 (VLA-5), одного из основных рецепторов фибронектина. Играет роль в межклеточных взаимодействиях.

Интегрин альфа-X — мембранный белок, гликопротеин из надсемейства интегринов, продукт гена ITGAX (CD11C), альфа-субъединица интегрина αXβ2, рецептора фибриногена. Ген был впервые клонирован в 1987 году.

Интегрин бета-6 (β6) — мембранный белок, гликопротеин из надсемейства интегринов, продукт гена ITGB6, бета-субъединица интегрина αVβ6, рецептора для фибронектина и цитотактина. Ген был впервые клонирован в 1990 году.

Синдекан 1 — мембранный белок, протеогликан семейства синдеканов, или трансмембранных гепарансульфатпротеогликанов. Всего описано 4 белка этого семейства. Продукт гена человека SDC1.

Синдекан 2, или гепарансульфатпротеогликан — мембранный белок, протеогликан семейства синдеканов, или трансмембранных гепарансульфатпротеогликанов. Продукт гена человека SDC2. Несёт на поверхности гепарансульфатные цепи.

EVI2B — мембранный белок, продукт гена EVI2B.
Терминальная оксидаза, или альтернативная оксидаза пластид, — фермент, который находится на стромальной стороне тилакоидной мембраны в хлоропластах растений и водорослей и внутриклеточных впячиваниях цитоплазматической мембраны цианобактерий. Впервые гипотеза о его существовании была высказан в 1982 году, и подтвердилась годами позже, после открытия в мембране тилакоида фермента, аминокислотная последовательность которого была сходной с таковой у альтернативной оксидазы митохондрий. Обе оксидазы произошли от общего предшественника — белка прокариот, и настолько схожи в структурном и функциональном плане, что помещённая в тилакоид альтернативная оксидаза может заменить выключенную терминальную оксидазу.
Суперсемейство белков — это наиболее крупная единица группировки белков, для которых может быть предположено наличие общего эволюционного предка. Обычно суперсемейство объединяется по принципу сходства вторичных и третичных структур белков, входящих в него и по принципу сходства механизма действия белков, несмотря на то, что сходства аминокислотных последовательностей в пределах суперсемейства может не наблюдаться и чаще всего не наблюдается. Суперсемейства обычно содержат несколько семейств белков, которые проявляют сходство аминокислотных последовательностей внутри семейств, но не между семействами. Термин «клан белков» часто применяется по отношению к суперсемействам протеаз, на основании классификации MEROPS.

Белки́, зая́коренные липи́дами, или липидосвя́занные белки́ — белки клеточной мембраны, ковалентно связанные с липидами клеточной мембраны. Эти липиды вставлены в мембрану бок о бок с хвостами жирных кислот. Белки, заякоренные липидами, могут находиться с любой стороны клеточной мембраны. Таким образом, липид служит своего рода якорем, закрепляющим белок вблизи клеточной мембраны.