Pararhabdodon
| † Pararhabdodon | |||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 Реконструкция | |||||||||||||||||||||||||||||||||||||||||
| Научная классификация | |||||||||||||||||||||||||||||||||||||||||
Домен: Царство: Подцарство: Без ранга: Без ранга: Тип: Подтип: Инфратип: Надкласс: Клада: Клада: Клада: Клада: Клада: Клада: Отряд: Подотряд: † Цераподы Инфраотряд: Парвотряд: Надсемейство: Семейство: Подсемейство: Триба: Род: † Pararhabdodon | |||||||||||||||||||||||||||||||||||||||||
| Международное научное название | |||||||||||||||||||||||||||||||||||||||||
| Pararhabdodon Casanovas-Cladellas et al., 1993 | |||||||||||||||||||||||||||||||||||||||||
| Синонимы | |||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||
| Единственный вид | |||||||||||||||||||||||||||||||||||||||||
† Pararhabdodon isonensis
| |||||||||||||||||||||||||||||||||||||||||
| Геохронология 70,6—66,0 млн лет
◄ Наше время◄ Мел-палеогеновое вымирание ◄ Триасовое вымирание◄ Массовоепермское вымирание ◄ Девонское вымирание◄ Ордовикско-силурийское вымирание◄ Кембрийский взрыв | |||||||||||||||||||||||||||||||||||||||||
Pararhabdodon (лат., возможное русское название — парарабдодон[3]) — род травоядных динозавров семейства гадрозаврид, живших в верхнемеловую эпоху (70,6—66,0 млн лет назад) на территории современной Испании[1]. Типовой и единственный вид Pararhabdodon isonensis назван и описан испанскими палеонтологами Casanovas-Cladellas, Santafe-Llopis и Isidro-Llorens в 1993 году. Родовое название означает «похожий на рабдодона», а видовое образовано от названия города Исона, недалеко от которого были найдены останки динозавра. В 1997 году Casanovas и колеги и Laurent и коллеги подкорректировали видовое название с isonense на isonensis, приведя в соответствие различия между родовым именем (Pararhabdodon — мужского рода) и видовым именем, данным от географического наименования (Исона — женского рода)[4].
История исследования
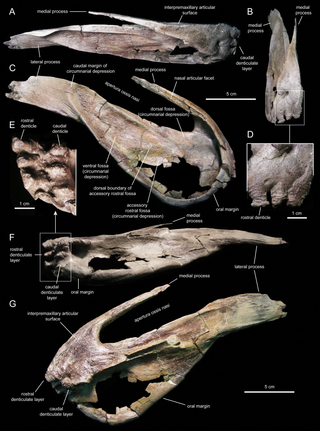
Голотип IPS SRA 1, паратипы и дополнительный материал найдены в слоях формации Тремп, датированных верхним маастрихтом, недалеко от города Исона, провинция Льейда, Испания. Голотип представляет собой почти полный средний хвостовой позвонок. Паратипы: IPS SRA 15 — левая плечевая кость, IPS SRA 16 — проксимальный фрагмент левой лопатки, IPS SRA 18 — шейные позвонки с краниального по срединный. Голотип и дополнительный материал, за исключением MCD 4919 (частичная левая челюстная кость), был собран в местонахождении Sant Roma d’Abella[5][4]; образец MCD 4919 был найден в местонахождении Serrat del Rostiar 1, недалеко от деревни Basturns. Sant Roma d’Abella моложе, чем Serrat del Rostiar 1, следовательно, образец MCD 4919 расширяет хроностратиграфический диапазон существования парарабдодона в верхнем маастрихте[6].

Материал включает останки черепа и нижней челюсти, части позвоночного столба, передних и задних конечностей. Все кости, за исключением челюсти, были собраны с площади 4×25 м, и, вероятно, принадлежат одной особи среднего размера (около 6 м длиной). Материал был обнаружен в период раскопок с весны 1985 по 1990 год. В работе Casanovas и коллег 1987 года материал (шейный и несколько частичных спинных позвонков, почти полная плечевая кость и фрагментарная лопатка) сначала определили, как родственный роду рабдодон. Голотип и паратипы описаны в 1993 году, как принадлежащие новому виду игуанодонтов — Pararhabdodon isonense. В 1994 году из того же местонахождения были извлечены две челюстные кости, два спинных позвонка, полный крестцовый отдел позвоночника, фрагмент седалищной кости и два фрагмента рёбер (образцы IPS SRA-19, 20, 22, 23, 24, 26)[5][4].
В местонахождении Les Llaus (у города Abella de la Conca), рядом с местонахождением Sant Roma d’Abella, на том же стратиграфическом горизонте, что и другие кости, была обнаружена правая зубная кость (IPS SRA 27). На основе стратиграфии, формы и размера зубная кость была отнесена как принадлежащая парарабдодону[5]. Однако Прието-Маркес с коллегами отвергли отнесение зубной кости к парарабдодону и в 2006 году на её основе описали новый вид гадрозавридов — Koutalisaurus kohlerorum. Хотя, подчеркнули, что, возможно, парарабдодон и Koutalisaurus являются синонимами[4]. В 2009 году Прието-Маркес с коллегами предложили новое филогенетическое положение парарабдодона, обнаружив поразительные сходства в морфологии с цинтаозавром. Что касается Koutalisaurus, то зубные кости циньтаозавра оказались очень похожи на зубную кость IPS SRA 27, отнесённую к Koutalisaurus, поэтому авторы предположили, что Koutalisaurus может являться младшим синонимом цинтаозавра (довод против — большое временное и пространственное разделение двух таксонов), либо парарабдодона (более вероятно). Однако перекрывающий материал пока не найден, чтобы подтвердить это наверняка[7].
Описание

Парарабдодон отличается от других гадрозаврид, за исключением цинтаозавра, высотой скуловой фасетки челюстной кости так, что вентральная часть располагается намного выше уровня латерального края эктоптеригоидного выступа, челюстная кость образует острый изгиб, простирающийся вентрально к скуловому отростку между скуловой фасеткой и эктоптеригоидным выступом; скуловая фасетка челюстной кости укорочена в передне-заднем направлении, вероятно, передняя часть скуловой кости узкая в передне-заднем направлении; передняя часть зубной кости с симфизным отростком, выступающим медиально так, что расстояние между симфизом и латеральной поверхностью зубной кости равняется толщине трёх губно-язычных альвеолярных полостей. От цинтаозавра парарабдодон отличается широкой, округлённо четырёхугольной переднедорсальной частью челюстной кости[7].
Филогения
Первоначально в 1993 году парарабдодон был описан (главным образом, на основе позвонков) как базальный игуанодонтид, близкий к рабдодону. Однако обнаруженные позднее кости (главным образом черепные и челюстные элементы) указали на однозначную близость к гадрозавридам. Согласно исследованию Casanovas и коллег 1999 года, парарабдодон является членом подсемейства ламбеозаврин на основе признаков: усечённое округлое сочленение челюстной кости со скуловой; каудальный конец челюстной кости усечён; вентрально отклонённая передняя часть зубной кости; очень высокие остистые отростки; дельтопекторальный гребень плечевой кости сильно выступает дистально. Более того, парарабдодон является базальным ламбеозаврином по наличию каудально рельефного крестца и умеренно расширенной седалищной кости[5]. Однако оба признака являются предковыми для гадрозавров, а рельефный крестец (гемальные борозды) проявляет внутривидовую изменчивость у некоторых таксонов[7]. Усечённое или округлое сочленение челюстной кости со скуловой не продемонстрировано, как синапоморфия ламбеозавринов (у Вейсхампеля и коллег, 1993, усечённый, закруглённый ростральный отросток скуловой кости рассматривается, как синапоморфия ламбеозавринов, но скуловая кость не известна у парарабдодона, и авторы не показывают, как этот признак отражается на морфологии челюстной кости). Заострённый дельтопекторальный гребень — примитивный признак, присутствующий у игуанодонтов. В дополнение, число максиллярных и дентарных зубов (около 35 в каждой кости) меньше, чем имелось у гадрозавринов и ламбеозавринов. Всё это говорит о том, что парарабдодон, вероятно, является базальным таксоном по отношению к последним[8].
В 2006 году Прието-Маркес с коллегами провели филогенетический анализ с использованием 127 признаков (93 краниальных и 34 посткраниальных) для 24 таксонов. Было получено 8 наиболее экономных деревьев. Во всех них парарабдодон находится внутри гадрозавроидов, но вне гадрозавридов. С гадрозавридами парарабдодон образовывал кладу[4].
В 2009 году Прието-Маркес с коллегами провёл новый анализ для 51 таксона. По его результатам парарабдодон и цинтаозавр формировали кладу внутри ламбеозаврин на основе следующих однозначных синапоморфий: длинный срединный выступ симфизной части зубной кости; почти прямая передняя беззубая часть зубной кости, служащая для сочленения с предзубной костью; приподнятое в дорсальном направлении сочленение челюстной кости со скуловой. Включение в анализ Koutalisaurus kohlerorum приводило к тому же результату, только парарабдодон, цинтаозавр и Koutalisaurus образовывали неразрешимую трихотомию. Включение парарабдодона в ламбеозаврины поддерживается следующими признаками: отсутствие максиллярного ростромедиального отростка; высокий дорсальный максиллярный отросток; соотношение между длиной диастемы зубной кости и зубной батареи составляет 0,2—0,31[7].
Кладограмма на основе филогенетического анализа Прието-Маркеса и коллег 2013 года[6]:
| Lambeosaurinae |
| ||||||||||||||||||||||||||||||||||||
Палеобиогеография
Возникновение в Европе в конце мелового периода примитивных ламбеозаврин, таких как парарабдодон, может быть объяснено с точки зрения географической изоляции. Из-за трансгрессии в течение альба — маастрихта Европа превратилась в архипелаг изолированных друг от друга фаунистических провинций. Продолжительность призрачной линии между возникновением первого ламбеозаврина и появлением парарабдодона составляет около 30 миллионов лет. Поэтому базальное расположение парарабдодона внутри ламбеозаврин и позднее стратиграфическое размещение таксона может быть объяснено викариансом[5].
Европейский архипелаг состоял из Английского, Иберо-Армориканского, Рейнско-Богемского, Адриатического, Австро-Альпийского и Трансильванского (остров Тисия-Дакия или Хатег) и других островов. У разных авторов разные реконструкции количества и площадей этих островов. В этом палеогеографическом контексте европейская гадрозавриновая фауна позднего маастрихта характеризовалась ламбеозавринами (аренизавр, бласизавр, парарабдодон и канардия из Иберо-Армориканского острова) и негадрозавридовыми гадрозавроидами. Тем не менее, на сегодняшний момент в Европе нет домаастрихтских останков, однозначно относимых к ламбеозавринам или гадрозавридам.
Statistical Dispersal-Vicariance анализ выводит Евразиатскую предковую область для самых последних общих предков Tsintaosaurini. Это должно значить, что расхождение парарабдодона от цинтаозавра представляет собой викарийные события, произошедшие не позднее раннего кампана. Вышеприведённый сценарий викарианса контрастирует с результатами Bayesian Binary MCMC и Dispersal Extinction Cladogenesis анализами. Оба метода выводят Азию как наиболее вероятный предковый ареал для цинтаозаврин. Согласно этой реконструкции происхождение парарабдодона должно явиться результатом расселения из Азии на запад Европейского архипелага. Викариантный сценарий, предложенный S-DIVA, находится в противоречии с текущими ископаемыми европейских ламбеозавринов. Как указывалось выше, не имеется окаменелостей ламбеозавринов, однозначно датированных временем до верхнего маастрихта.
Поэтому предковые арелы, полученные посредством анализов BBM и DEC и соответствующие сценариям расселения, больше согласуются с известными ископаемыми ламбеозавринов в Европе. Tsintaosaurini, по всей видимости, достигли Иберо-Армориканского острова в конце раннего или в позднем маастрихте. Биогеографические сценарии, включающие расселение ламбеозавринов из Азии на Европейский архипелаг, ранее предлагались различными авторами, хотя указывали время расселения — середину верхнего кампана, что раньше, чем оценка, основанная на самых ранних появлениях окаменелостей этих гадрозавридов в слоях Европы[6].
Примечания
- ↑ 1 2 †Pararhabdodon (англ.) информация на сайте Fossilworks. (Дата обращения: 2 апреля 2016)
- ↑ †Pararhabdodon isonensis (англ.) информация на сайте Fossilworks. (Дата обращения: 2 апреля 2016)
- ↑ Циньтаозавр Архивная копия от 18 апреля 2016 на Wayback Machine на сайте Мир природы
- ↑ 1 2 3 4 5 Prieto-Marquez, A., Gaete, R., Rivas, G., Galobart, A., and Boada, M. (2006). Hadrosauroid dinosaurs from the Late Cretaceous of Spain: Pararhabdodon isonensis revisited and Koutalisaurus kohlerorum, gen. et sp. nov. Journal of Vertebrate Paleontology 26(4): 929—943.
- ↑ 1 2 3 4 5 Casanovas, M.L, Pereda-Suberbiola, X., Santafe, J.V., and Weishampel, D.B. (1999). First lambeosaurine hadrosaurid from Europe: palaeobiogeographical implications. Geological Magazine 136(2): 205—211.
- ↑ 1 2 3 A. Prieto-Marquez, F. M. D. Vecchia, R. Gaete and A. Galobart. (2013). Diversity, relationships, and biogeography of the Lambeosaurine dinosaurs from the European archipelago, with description of the new aralosaurin Canardia garonnensis. PLoS One 8(7): e69835
- ↑ 1 2 3 4 Prieto-Marquez, A.; Wagner, J.R. (2009). Pararhabdodon isonensis and Tsintaosaurus spinorhinus: a new clade of lambeosaurine hadrosaurids from Eurasia. Cretaceous Research. online preprint (5): 1238.
- ↑ Head, J.J. (2001). A reanalysis of the phylogenetic position of Eolambia caroljonesa (Dinosauria: Iguanodontia). Journal of Vertebrate Paleontology 21: 392—396.