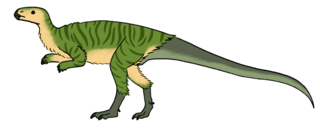
Pisanosaurus
| † Pisanosaurus | |
|---|---|
 Реконструкция черепа. Белым цветом обозначены сохранившиеся кости; светло-серым цветом обозначены кости, известные по сильно размытым отпечаткам | |
| Научная классификация | |
Домен: Царство: Подцарство: Без ранга: Без ранга: Тип: Подтип: Инфратип: Надкласс: Клада: Клада: Клада: Клада: Клада: Клада: Отряд: Род: † Pisanosaurus | |
| Международное научное название | |
| Pisanosaurus Casamiquela[англ.], 1967 | |
| Единственный вид | |
Писанозавр[1] (лат. Pisanosaurus) — род примитивных растительноядных динозавроподобных пресмыкающихся из позднего триаса Южной Америки. Ранее считался одним из самых ранних представителей птицетазовых динозавров. Типовой и единственный вид Pisanosaurus mertii назван и описан аргентинским палеонтологом Родольфо М. Касамикела (Rodolfo M. Casamiquela) в 1967 году. Родовое имя «ящер Писано» названо в честь аргентинского палеонтолога Хуан Писано (Juan Pisano). Видовое имя названо в честь арауканского натуралиста Карлос Мерти (Carlos Merti)[2].
История исследования
Голотип PVL 2577 обнаружен Galileo J. Scaglia в слое формации Исчигуалисто, датированным карнием триаса, 237—227 млн лет назад, провинция Ла-Риоха, Аргентина. Эрозия уничтожила большую часть черепа, левой части нижней челюсти, большую часть передних конечностей, таза и хвоста. Изначально экземпляр состоял из 21 сочленённого позвонка, прикреплённых к черепу, и сильно пострадавших от эрозии. Casamiquela в своей работе описал большинство из доступных останков писанозавра, за исключением отпечатков таза, тел крестцовых позвонков и трёх пястных костей. Он ошибочно определил шейные позвонки, как поясничные и хвостовые.
Голотип включает: фрагмент правой верхнечелюстной кости с 11 плотно упакованными зубами в альвеолах, неполную часть правой нижней челюсти (без предзубной кости) с 15 плотно упакованными зубами (пространство между зубным рядом и краем зубной кости довольно широкое), неопределённый фрагмент левой части нижней челюсти, 6 неполных шейных позвонков, 7 неполных грудных позвонков, отпечатки 5 крестцовых позвонков, ребро и пара фрагментов, возможно, относящихся к рёбрам, неполную правую лопатку, отпечатки трёх пястных костей, неполный отпечаток правой половины таза, неполные, плохо сохранившиеся обе бедренные кости, правые большеберцовую и малоберцовую кости, сочленённые с таранной и пяточной костями, маленькую вторую заплюсневую дорсальную кость, соединённую с плюсневой, третью и четвёртую плюсневые кости с сочленёнными фалангами пальцев, неопределённый фрагмент длинной кости[2][3].
Пол Серено сомневается, что отпечатки 5 тел позвонков — это отпечатки именно крестцовых позвонков. Передние три тела не сильно сочленены между собой; нет свидетельств наличия крестцовых рёбер вблизи их сторон. Это скорее всего задние грудные позвонки. Задние два тела прочно соединены друг с другом и первое имеет латеральный выступ для крепления крестцового ребра. Отпечаток ребра присутствует недалеко от седалищного стебля подвздошной кости. Число крестцовых позвонков не может быть определено[4].
Irmis et al., 2007, высказывают сомнения, что Bonaparte правильно определил шейные позвонки, которые Casamiquela считал хвостовыми. Авторы оставляют их неопределёнными, хотя морфология напоминает всё-таки хвостовые[5].
Описание

Небольшое, двуногое динозавроподобное пресмыкающееся, хороший бегун. Отличается от других известных форм псевдозухий, птицетазовых и ящеротазовых динозавров рядом комбинаций общих и специализированных признаков, среди которых: наличие ямки аддуктора на нижней челюсти, отношение длины задентарной части по отношению к дентарию, морфология позвоночника и задних конечностей. А также низкое положение мыщелка нижней челюсти, морфология зубной кости (с крепким основанием), однородность и распределение частокола зубов в дополнении к форме окклюзии[2].
Точно не известно, сколько зубов располагалось в верхней челюсти, но сравнение с нижней челюстью даёт предположить, что их было 15—16. Длина бедренной кости предположительно равна длине или немного короче большеберцовой + таранной[3]. Дистальный конец большеберцовой кости почти квадратный. В отличие от большинства птицетазовых, толщина дистального конца превышает его максимальную ширину. Скорее всего, эта пропорция является аутапоморфией у писанозавра. Очень толстый восходящий отросток таранной кости является плезиоморфией в сравнении с птицетазовыми и ящеротазовыми; его толщина составляет больше половины толщины таранной кости. Дискообразная пяточная кость является аутапоморфией[4].
Длина Pisanosaurus оценивается Грегори Полом в 1,3 м, а вес в 2 кг[6].
Филогения
По результатам кладистического анализа Butler et al., 2008, с использованием 221 признака для 46 таксонов, поддерживается позиция писанозавра, как самого базального таксона птицетазовых, но не исключается возможность принадлежности писанозавра к гетеродонтозавридам[7].
Кладограмма по результатам анализа Butler et al., 2008:
| Ornithischia |
| ||||||||||||||||||||||||
Филогенетическое положение Pisanosaurus крайне проблематичное. Casamiquela поместил писанозавра к орнитоподам, как очень примитивную форму, введя для него новое семейство Pisanosauridae[2].
Richard A. Thulborn в своих работах отмечал тесное родство писанозавра с фаброзавром. Оба рода при этом входили в семейство гипсилофодонтидов[8][9]. Палеонтолог Петер М. Гальтон в 1972 году, напротив, не находит больших сходств писанозавра с фаброзавром и относит первого к очень примитивным гипсилофодонтидам, от похожих форм которого, возможно, произошли все остальные представители семейства, а фаброзавра выводит за пределы гипсилофодонтид, вводя для него новое семейство Fabrosauridae[10]. Ныне клада Fabrosauridae вышла из употребления, т. к. с 1991 года фаброзавру присвоен статус nomen dubium из-за недостаточности ископаемого материала.
Несмотря на отсутствие гетеродонтии у писанозавра Jose F. Bonaparte находит сходство в морфологии и износе верхних и нижних «заклыковых» зубов у гетеродонтозавра и писанозавра: зубы почти цилиндрические, плотно упакованы друг с другом и в процессе износа образуется плоская или слегка вогнутая жевательная поверхность, образующая более-менее непрерывную плоскость вдоль всего ряда зубов. Однако у гетеродонтозавра имелись вертикальные гребни на дистальной стороне зубов, отсутствующие у писанозавра, хотя, возможно, это результат сильного износа зубов. Отсутствие гребней и гетеродонтии Bonaparte считает недостаточным основанием для разделения писанозавра и гетеродонтозавра по разным семействам. Касательно гипсилофодонтидов, куда Galton поместил писанозавра, Bonaparte отмечает, что коренные зубы гипсилофодона листовидной формы, сжатые с боков, с гребнями на внутренней поверхности нижних и внешней поверхности верхних зубов; также у него имеется непрерывная смена зубов. Все эти признаки отсутствуют у писанозавра. Хотя, по поводу смены зубов есть некоторая неопределённость, поскольку имеется скелет только одной особи. Морфология зубов фаброзавра и писанозавра также сильно разниться. Подводя итог, Bonaparte помещает писанозавра в семейство гетеродонтозавридов, которое имеет приоритет над семейством Pisanosauridae[3]. В настоящее время семейство Pisanosauridae не считается валидным и авторами не используется[11].
Пол Серено отмечает, что фрагменты челюсти, частичный отпечаток таза и дистальная правая задняя конечность могут принадлежать одной особи, но фрагментарная лопатка и другие части посткраниального скелета могут быть частями другого животного, потому что пропорционально слишком маленькие. Щёчная выемка более продвинутая, чем у лесотозавра. Коронки дентарных и максиллярных зубов имеют цингулюм и находятся под углом к корням, как у многих птицетазовых. Износ граней сильно уменьшает коронки, усекая дистальные верхушки. Углы граней износа зубов немного разнятся, а смежные края коронок не выравнены друг относительно друга. Следовательно, грани износа не образуют сплошную жевательную поверхность, как у гетеродонтозавра, что противоречит выводам Bonaparte. Таз динозавра не демонстрирует каких-то птицетазовых модификаций и примитивен в этом плане. Лобковая кость выступает антеровентрально и имеет глубокий дорсовентральный контакт с седалищной костью, чего не наблюдается у птицетазовых. Серено резюмирует: если череп и посткраниальная часть голотипа верно ассоциированы, то писанозавр, возможно, является сестринским таксоном к другим птицетазовым[4].
Irmis et al., 2007, уверенно отклонили предположение Серено, что голотип писанозавра может состоять из образцов двух особей или таксонов. Наличие венечного отростка зубной кости является синапоморфией птицетазовых. Посткраниальный скелет не сохранил синапоморфий динозавров. Проксимальный конец большеберцовой кости — это плезиоморфный признак и похож на таковые у базальных Dinosauriformes: Silesaurus, Pseudolagosuchus и Marasuchus. Сочетание птицетазовых черепно-зубных признаков, более продвинутых, чем у лесотозавра, и посткраниальных плезиоморфных признаков динозавроподобных затрудняет определение филогенетического положения писанозавра. Поскольку все признаки, выводящие динозавра за пределы птицетазовых, являются плезиоморфиями, писанозавр рассматривается авторами, как птицетазовый динозавр. Тем не менее, поскольку все синапоморфии птицетазовых связаны с питанием, гипотеза, что писанозавр является базальным Dinosauriformes с челюстной морфологией конвергентно сходной с птицетазовыми, не могла быть исключена и нуждалась в строгом филогенетическом анализе. Проблема предыдущих анализов в том, что в них писанозавр считался динозавром априори. В новый анализ должны были быть включены нединозавровые внешние группы, базальные динозавроподобные, базальные динозавры и разнообразные виды птицетазовых[5].
Филогенетический анализ, проведенный Agnolin в 2015 году, выявил писанозавра как возможного нединозаврового члена Dinosauriformes, относящегося к силезавридам.[12] В 2017 году вновь было выдвинуто предположение, что писанозавр был силезавридом.[13][14]
Точная классификация Pisanosaurus была темой обсуждения уже более 40 лет; До 2017 года считалось, что писанозавр является самым первым из известных орнитисхий, разнообразной группы динозавров жившей в течение почти всей мезозойской эры, но сейчас некоторые учёные считают, что писанозавр это нединозавровый силезаврид.[13][14][15]
Палеоэкология
Формация Исчигуалисто была вулканически активной поймой, покрытой лесами, с тёплым и влажным климатом,[16] хотя была подвержена сезонным колебаниям, включая обильные дожди.[17] Растительность формации состояла из папоротников, хвощей и гигантских хвойных деревьев, которые образовывали высокогорные леса вдоль берегов рек.[18] Травоядные были представлены ринхозаврами, такими как гиперодапедон, примитивными этозавровыми текодонтами как Aetosaurus, каннемейериидами, такими как Ischigualastia и Exaeretodon, и траверсодонтидами (Traversodontidae). Эти нединозавровые травоядные были намного более многочисленными, чем ранние динозавры.[19] Рядом обитали плотоядные престозухиды (Prestosuchidae), примитивные ящеротазовые, орнитозухиды, хищные и гомфодонтные цинодонты и другие травоядные и плотоядные животные, хронологический период существования которых плохо определён[3]. Похоже, что остатки герреразавра были наиболее распространёнными среди плотоядных животных формации Исчигуалисто.[20]
Писанозавр обитал в густых хвойных лесах, обильно поливаемых дождями. Его соседями были травоядный Panphagia, а также хищники Eoraptor и Herrerasaurus[6].
Примечания
- ↑ Барон М. (ред.). Динозавры. Полный иллюстрированный словарь = Dictionary of Dinosaurs: An Illustrated A to Z of every Dinosaur Ever Discovered : ориг. изд. 2018 : [пер. с англ.]. — М. : Эксмо, 2021. — С. 121. — 184 с. — ISBN 978-5-04-115355-7.
- ↑ 1 2 3 4 Casamiquela, R.M. (1967). «Un nuevo dinosaurio ornitisquio triasico (Pisanosaurus mertii; Ornithopoda) de la Formacion Ischigualasto, Argentina». Ameghiniana 4 (2): 47—64.
- ↑ 1 2 3 4 Bonaparte, J.F. (1976). «Pisanosaurus mertii Casamiquela and the origin of the Ornithischia». Journal of Paleontology 50 (5): 808—820.
- ↑ 1 2 3 Sereno, P.C. (1991). «Lesothosaurus, „fabrosaurids“, and the early evolution of Ornithischia». Journal of Vertebrate Paleontology 11 (2): 168—197.
- ↑ 1 2 Irmis, R.B.; Parker, W.G.; Nesbitt, S.J.; Liu, J. (2007). «Early ornithischian dinosaurs: the Triassic record». Historical Biology 19: 3—22.
- ↑ 1 2 Paul, G.S., 2010, The Princeton Field Guide to Dinosaurs, Princeton University Press p. 214.
- ↑ Butler, Richard J.; Upchurch, Paul and Norman; David, B. (2008). The phylogeny of the ornithischian dinosaurs. Journal of Systematic Palaeontology 6 (1): 1—40.
- ↑ R. A. Thulborn. 1970. The skull of Fabrosaurus australis, a Triassic ornithischian dinosaur. Palaeontology 13(3):414-432.
- ↑ R. A. Thulborn. 1972. The post-cranial skeleton of the Triassic ornithischian dinosaur Fabrosaurus australis. Palaeontology 15(1):29-60.
- ↑ P. M. Galton. 1972. Classification and evolution of ornithopod dinosaurs. Nature 239:464—466.
- ↑ Taxon Search : View Taxon Details. Дата обращения: 29 октября 2015. Архивировано из оригинала 6 марта 2016 года.
- ↑ Federico L. Agnolin (2015). "Nuevas observaciones sobre Pisanosaurus mertiiCasamiquela, 1967 (Dinosauriformes) y sus implicancias taxonómicas" (PDF). XXIX Jornadas Argentinas de Paleontología de Vertebrados. 27–29 de Mayo de 2015. Diamante, Entre Ríos. Libro de Resúmenes: 13–14. Archived from the original (PDF) on 2016-01-27. Retrieved 2015-08-07.
- ↑ 1 2 Agnolín, Federico L.; Rozadilla, Sebastián (2017). "Phylogenetic reassessment of Pisanosaurus mertii Casamiquela, 1967, a basal dinosauriform from the Late Triassic of Argentina". Journal of Systematic Palaeontology. 16 (10): 853–879. doi:10.1080/14772019.2017.1352623.
- ↑ 1 2 Matthew G. Baron; David B. Norman; Paul M. Barrett. Baron et al. reply (англ.) // Nature. — 2017. — Vol. 551, no. 7678. — P. E4–E5. — doi:10.1038/nature24012. — . — PMID 29094705.
- ↑ Matthew G. Baron (2018). "Pisanosaurus mertiiand the Triassic ornithischian crisis: could phylogeny offer a solution?". Historical Biology: An International Journal of Paleobiology. 31 (8): 1–15. doi:10.1080/08912963.2017.1410705.
- ↑ Tucker, Maurice E.; Benton, Michael J. (1982). "Triassic environments, climates, and reptile evolution" (PDF). Palaeogeography, Palaeoclimatology, Palaeoecology. 40 (4): 361–379. Bibcode:1982PPP....40..361T. doi:10.1016/0031-0182(82)90034-7. Archived from the original (PDF) on 2009-01-26. Retrieved 2009-07-23.
- ↑ Columbi, Carina E. (2008-10-05). Stable isotope analysis of fossil plants from the Upper Triassic Ischigualasto Formation in the northwest of Argentina. Houston, TX: The Geological Society of America. Archived from the original on 2012-01-11. Retrieved 2009-07-23.
- ↑ Sereno, P.C.; Novas, F.E. (1992). "The complete skull and skeleton of an early dinosaur". Science. 258 (5085): 1137–1140. Bibcode:1992Sci...258.1137S. doi:10.1126/science.258.5085.1137. PMID 17789086.
- ↑ Bonaparte, J.F. (1970). "Annotated list of the South American Triassic tetrapods". Gondwana Symposium Proceedings and Papers. 2: 665–682.
- ↑ Rogers, R. R.; Swisher III, C.C.; Sereno, P.C.; Monetta, A.M.; Forster, C.A.; Martinez, R.N. (1993). "The Ischigualasto tetrapod assemblage (Late Triassic, Argentina) and 40Ar/39Ar dating of dinosaur origins". Science. 260 (5109): 794–797. Bibcode:1993Sci...260..794R. doi:10.1126/science.260.5109.794. PMID 17746113.
| Таксономия |
|---|